
Tags
A-Group, Afro-Asiatic, Ala111Thr, ancient DNA, Ancient Egypt, Badarian, biogeographical analysis, Brenda Stoessiger, C-Group, Capsian, Carleton Coon, Cushitic, Daniel Stiles, EDAR, Ethiopia, G. P. Rightmire, Hamitic, Harold C. Fleming, Iberomaurusian, Indus Valley, Kiffian, lactose tolerance, Linearbandkeramik, Mediterranean, Meroe, Mota, Nella Puccioni, Omotic, Out-of-Africa, Pastoral Neolithic, Richard Leakey, Robert Gayre, Samuel George Morton, Savanna Pastoral Neolithic, Semitic
Scientists often regard Ethiopia as the cradle of humanity. In the 1960s, an archaeological expedition under Richard Leakey discovered skeletal remains at the Omo-Kibish sites near the Omo river. Since these fossils show some modern features and have been tentatively dated to almost 200,000 years before present, many paleontologists argue that they represent the earliest anatomically modern humans (Homo sapiens sapiens) found so far. Other scholars dispute this association, remarking that the skulls also possess archaic characteristics; particularly the Omo II specimen, which has retained the robusticity of earlier, non-modern hominids.

Various evolutionary theories on the origins and dispersal of modern humans (Groucutt et al. (2015)).
The “Out-of-Africa” (OOA) or “Recent African Origin” (RAO) theory emerged as one of several competing hypotheses seeking to explain the prehistoric peopling of the world. Although initially born out of a hoax, complete with false dating, the OOA model gained popularity in the 1990s with the development of the human paternal (Y-DNA) and maternal (mtDNA/mitochondrial DNA) phylogenetic trees. Researchers observed that the deepest or oldest uniparental lineages on the respective trees, paternal haplogroup A and maternal haplogroup L0, are mainly restricted to small Khoisan groups inhabiting Southern Africa. They also found that most populations outside of Africa carry younger maternal lineages that were prehistorically derived from the L3 macroclade (“Eurasian Eve”). Although widely diffused both within and outside of Africa, this haplogroup today has its greatest diversity in Ethiopia. Consequently, L3 is assumed to have been in the area for at least several millenia i.e. long enough to have evolved various sublineages, and thus likely to have originated in the region. Ethiopia is therefore often considered the most probable starting point of the suggested Out-of-Africa colonization.
For these and other reasons, ancient DNA from human fossils in the area holds the potential to greatly improve our understanding of global prehistory.
Mota
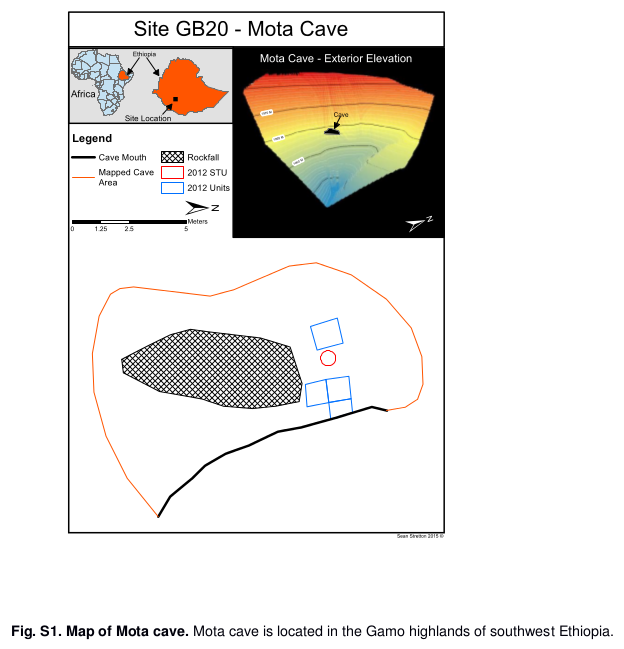
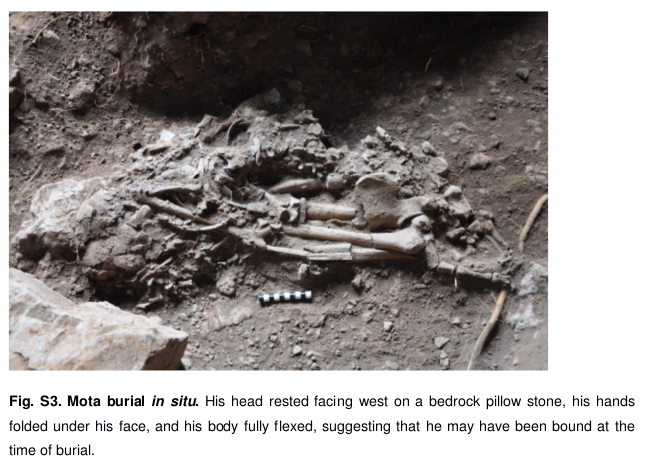
In 2015, a genetic research team led by M. Gallego Llorente and E. R. Jones managed to successfully extract ancient DNA from a human skeleton found in Mota Cave, located in the Gamo highlands of southwestern Ethiopia. The Mota remains belonged to a middle-aged male hunter-gatherer, and were radiocarbon dated to around 4,500 years before present:
 Low contamination rates were observed, confirming the authenticity of the extracted DNA. Further examination of the fossil’s Y-DNA and mtDNA assigned him to the paternal haplogroup E1b1 and the maternal haplogroup L3x2a, respectively:
Low contamination rates were observed, confirming the authenticity of the extracted DNA. Further examination of the fossil’s Y-DNA and mtDNA assigned him to the paternal haplogroup E1b1 and the maternal haplogroup L3x2a, respectively:
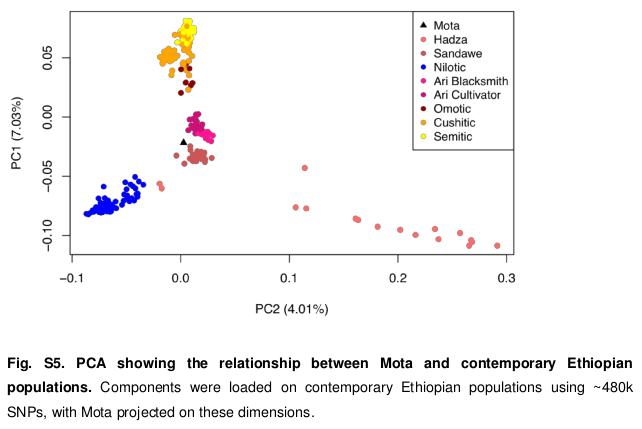
 To more closely gauge Mota’s population affinities, the scientists also ran a principal component analysis comparing his DNA against that of various contemporary Ethiopian populations from different ethnolinguistic groups. The specimen was most similar to the South Omotic-speaking Ari and the Khoisan-speaking Sandawe populations:
To more closely gauge Mota’s population affinities, the scientists also ran a principal component analysis comparing his DNA against that of various contemporary Ethiopian populations from different ethnolinguistic groups. The specimen was most similar to the South Omotic-speaking Ari and the Khoisan-speaking Sandawe populations:
 The close association between Mota and the modern Ari was also supported by f3 statistical analysis, which showed that they formed a clade unto themselves.
The close association between Mota and the modern Ari was also supported by f3 statistical analysis, which showed that they formed a clade unto themselves.
In order to ascertain whether Mota harbored any West Eurasian ancestry like modern Ethiopian populations, the researchers ran an admixture analysis using the Yoruba and Druze as the African and West Eurasian reference samples, respectively, against which Mota’s DNA and that of other contemporary populations was compared. The results suggested that Mota lacked any West Eurasian ancestry. This was also supported by the fact that the specimen did not carry the derived SLC24A5 (Ala111Thr/rs1426654) allele linked with lighter skin pigmentation, nor any lactase persistence variants, nor apparently any Neanderthal-associated alleles.
So what can be concluded from this ancient DNA analysis? Are the findings significant, or is Mota simply an early Ari individual and little beyond that?
The results are quite illuminating in that they suggest, among other things, that:
- Mota’s nearest contemporary relatives are the Ari.
- A biological division exists between speakers of the South Omotic languages and other local Afro-Asiatic-speaking populations (including North Omotic speakers).
- Mota co-existed in the region with physically and genetically distinct early Cushitic populations. Hodgson et al. (2014)’s inferred “Ethio-Somali” (ancestral Afro-Asiatic) and “Ethiopic” (ancestral Ari) components are therefore real.
- There also appears to be as yet unidentified East Eurasian ancestry in the Horn, and likely elsewhere in Northeast Africa.
- Using ancient DNA samples as reference groups in lieu of modern samples can potentially uncover hitherto obscured population affinities.
- The prehistoric Out-of-Africa colonization, if any, probably did not emanate from Ethiopia.
Limitations of biogeographical analysis
Before proceeding further, let us briefly note some of the principal limitations of biogeographical analysis, such as that around which the Mota study is centered. BGA/admixture testing is at its core speculative, for it is based on probability. That is, such analysis estimates where or in which geographical area given stretches of DNA known as SNPs likely originated according to which reference populations today carry those SNPs at highest frequencies. This is problematic for a number of reasons, some quite obvious:
- BGA testing is entirely dependent on the quality of the modern reference populations that the SNPs are compared against. For example, if a modern reference population with multiple ancestries is included in the analysis (like, say, the Maasai, who are Nilotes with some Cushitic influence), the SNPs could theoretically be most closely associated with any of that mixed reference population’s various ancestries (in this case, they could be associated with either the early Nilotes or the early Cushites).
- Where a reference population resides today is not necessarily where it resided in the past.
- Just because a reference population today has the highest frequencies of a particular SNP does not necessarily mean it always did. For one thing, many ancient populations have died off (early Cushitic populations in particular) or gone through population bottlenecks, and they easily could have had higher frequencies of that SNP than any modern reference population.
- Just because a reference population today has the highest frequencies of a particular SNP does not necessarily mean that that SNP actually originated with that proxy group. This SNP could have been passed on to that reference population’s ancestors by interbreeding with an unrelated, now extinct population. Through linguistics and haplogroup analysis, we know for a fact that such contacts happened many times in the past and over a wide area between now extinct Cushitic groups and early Nilotic and Bantu populations.
- Certain modern populations have high frequencies of private alleles, which are genetic variants that are only found today in that population. These are quite common among contemporary Afro-Asiatic-speaking groups in Northeast Africa, who have among the most private alleles on the continent. In other words, many SNPs for these individuals cannot be matched to any modern reference population. They are therefore not factored into the biogeographical analysis, which in turn gives a misleading estimate as to where most of these individuals’ overall SNPs truly cluster.
- Ancient reference populations, though certainly preferable to modern proxy groups, are also subject to many of these same caveats; particularly if they have not been carefully selected. An example of this is shown below, where it is demonstrated that neither the Mota specimen nor the Neolithic LBK sample used in the Mota analysis are representative of the ancestral Afro-Asiatic speakers in the Horn. More appropriate ancient proxy groups are instead suggested (under final observations and recommendations).
Besides the points above, perhaps the biggest limitation of biogeographical analysis is the fact that it can only at best capture a small fraction of an individual’s total ancestry i.e. that contained within the genetic tree, not that within the exponentially larger genealogical tree. The Genetic Genealogist explains:
In reality, everyone has two family trees. The first is a Genealogical Tree, which is every ancestor in history that had a child who had a child who had a child that ultimately led to you. Every decision made by every person in that tree contributed to who and what you are today.
However, not every person in that tree contributed a segment of your DNA sequence (because of random inheritance, as discussed above). As a result, we have a second family tree – a Genetic Tree – which is a tree that contains only those ancestors who contributed to our DNA. No one has yet been able to construct their Genetic Tree, but soon it will be a reality thanks to advances in genetic sequencing and comparison such Relative Finder. These tools are using relatedness between people living today to deduce the inheritance of DNA from people who have been dead for centuries.[…]
The Genealogical Family Tree contains ALL of your biological ancestors[…] The Genetic Family Tree contains a small subset of your biological ancestors[…] Due to the nature of the Genealogical versus the Genetic Family Tree, entire populations, ancestors, and ethnicities are regularly lost entirely from your DNA! [They] therefore would not be detected by a DNA test.
Uniparental markers and Out-of-Africa
The Mota specimen’s paternal haplogroup E1b1 is today relatively rare, and is mainly restricted to a few Afro-Asiatic speakers in Ethiopia and environs. It has been found amongst 18% of Ethiopian Jews, 11%-12.8% of Oromos, 11% of Iraqw, 6%-10.4% of Amhara, 10% of Ethiosemitic speakers generally, and in 18.2% of Ethiopians as a whole. Given the clade’s close association with Afro-Asiatic-speaking populations in Ethiopia, it appears that Mota’s ancestors obtained the E1b1 haplogroup through contact with early Afro-Asiatic male settlers in the area.
Mota’s L3x2a maternal lineage points to a similar, if more ambiguous, affiliation. It too is today mainly concentrated among some Afro-Asiatic speakers in East Africa, and is also present in Egypt and among Yemeni Jews. How exactly Mota’s hunter-gatherer culture acquired the haplogroup is uncertain, for Babalini et al. (2002) found a comparably-aged L3 carrier in their ancient DNA study of early human specimens from the Fezzan in Libya (dated to around 3,000-1,500 BCE). Fernández et al. (2014) likewise identified an even older L3-bearing individual in their examination of skeletal remains from Pre-Pottery Neolithic B (PPNB) sites in the Near East. Dated to between 8,700-6,600 BCE, the PPNB specimen predates Mota and the Fezzan individual by over four thousand years. This highlights the early global distribution of the L3 haplogroup, if not an ultimately Middle Eastern origin for the clade, as Farrell et al. (2013) propose:
Here we present the first high-coverage whole genome sequences from a Middle Eastern population consisting of 14 Eastern Province Saudi Arabians. Genomes from this region are of interest to further answer questions regarding “Out-of-Africa” human migration. Applying a pairwise sequentially Markovian coalescent model (PSMC), we inferred the history of population sizes between 10,000 years and 1,000,000 years before present (YBP) for the Saudi genomes and an additional 11 high-coverage whole genome sequences from Africa, Asia and Europe.
The model estimated the initial separation from Africans at approximately 110,000 YBP. This intermediate population then underwent a long period of decreasing population size culminating in a bottleneck 50,000 YBP followed by an expansion into Asia and Europe. The split and subsequent bottleneck were thus two distinct events separated by a long intermediate period of genetic drift in the Middle East. The two most frequent mitochondria haplogroups (30% each) were the Middle Eastern U7a and the African L. The presence of the L haplogroup common in Africa was unexpected given the clustering of the Saudis with Europeans in the phylogenetic tree and suggests some recent African admixture. To examine this further, we performed formal tests for a history of admixture and found no evidence of African admixture in the Saudi after the split. Taken together, these analyses suggest that the L3 haplogroup found in the Saudi were present before the bottleneck 50,000 YBP. Given the TMRCA estimates for the L3 haplogroup of approximately 70,000 YBP and the timing of the Out-of-Africa split, these analyses suggest that L3 haplogroup arose in the Middle East with a subsequent back migration and expansion into Africa over the Horn-of-Africa during the lower sea levels found during the glacial period bottleneck.
These results are consistent with the hypothesis that modern humans populated the Middle East before a split 110,000 YBP, underwent genetic drift for 60,000 years before expanding to Asia and Europe as well as back-migration into Africa. Examination of genetic variants discovered by Saudi whole genome sequencing in ancestral African populations and European/Asian populations will contribute to the understanding human migration patterns and the origin of genetic variation in modern humans.
Thus, the prehistoric “Out-of-Africa” colonization, assuming one occurred at all, does not appear to have emanated from the Horn. The finding that the L3 haplogroup likely originated in and spread from the Middle East points instead to some other center of evolution. (*N.B. For the latest evidence on the suggested non-African origin of the mtDNA macrohaplogroup L3, see Cabrera (2022) and Cabrera et al. (2017).)
North Omotic vs. South Omotic

Kawo Tona, the last King of the Wolayta. Prior to assimilating Mota-related foragers, most Omotic speakers had a Cushitic physiognomy like that of this late ruler. Nowadays, this phenotype is mainly confined to North Omotic speakers.
The Omotic branch of Afro-Asiatic is divided into two subgroups: North (also known as Nomotic) and South (or Somotic). South Omotic consists of the Ari, Dime, Hamar, Gayil and Karo languages. Of these South Omotic idioms, the first three are sometimes collectively known as “Aroid”.
Anthropologists and linguists working in the Ethiopian region have long observed a marked physical and linguistic cleavage between, on the one hand, the speakers of the Aroid Omotic languages, and on the other, the non-Aroid Afro-Asiatic-speaking populations. Harold C. Fleming, who coined the term “Omotic” and helped establish the validity of the phylum as an independent branch of the Afro-Asiatic family, remarks that the Ari are generally “Negroid” in physiognomy, in contrast to the “Ethiopid” northern Cushitic and Ethiosemitic groups:
The Ari peoples have been described extensively by scholars of the Frobenius tradition, especially A. Jensen, Eike Haberland, and others, as well as observed and described more informally by various anthropologists (e.g., Herbert Lewis, Jean Lydall, Ivo Strecker, myself) and Ethiopian government officials. One of our colleagues, Ayyalew Mitiku, is an Amhara who grew up among the Ari and speaks their language fluently. Among all these observers there is a consensus that the Ari and many of the Banna and Hamar represent a variety of African Negro, rather than an Ethiopid or “Afro-Mediterranean” variety. However, the Ari variety of Negro is distinctive and is not very much like the neighbouring Nilotic or Surmic peoples to the west and south; nor particularly like the Bantu of East Africa. Some resemblance is noted to the Koman of the Ethiopia-Sudan border areas and some Sudanese populations. Above all this aspect of the Ari, while it has earned them the label of “blacks” or “Shanqillas”, strikes many of us as being the appearance of a distinctive kind of African, a population that has evolved on its own to a considerable extent.
Due to the Ari’s aberrant morphology and the divergent nature of the Aroid languages themselves, Fleming further indicates that Ari and the other Aroid languages were regarded by earlier scholars as non-Afro-Asiatic idioms possessing Nilo-Saharan affinities:
The three main branches of my Somotic, viz, Dime, Hamar and the Ari cluster are what [Lionel Bender] calls Aroid; they are so different from the rest of Omotic that older German and Italian scholars thought of them as ‘Negroid’ or Nilotic or the like. It was a struggle to get them accepted as a branch of Omotic or West Cushitic. All you have to do is work on a Dizoid (Maji, Na’o, Shako) language and then on Dime or Galila (Ari), as I did, to see that they are very different in phonology and morphology. Yes, there has been some borrowing across the Omo river. Dime informants will tell you that “Maji” used to rule them; the borrowings are not massive, however. Dizoid and ‘Aroid’ simply do not belong in the same moiety in opposition to the rest of Omotic.
Correspondingly, in the ethnological literature, the traditional explanation for the Ari’s differing physiognomy and culture as well as the divergent nature of their South Omotic language is that they are descendants of Nilo-Saharan-speaking peoples (the Pre-Nilotes), who settled in the parts of Ethiopia bordering South Sudan around 4,000 years ago. These Pre-Nilote groups are then thought to have interbred with early Omotic populations from the Sahara and gradually adopted the latter’s Afro-Asiatic languages, thereby giving rise to the Ari and related groups. Per the Centre National de la Recherche Scientifique:
Negroid people from the Sudan, speaking languages ancestral to those classified as the four branches of the Nilo-Saharan family settled in West and South West Ethiopia about the third millenium B.C. (Pre-Nilotes). They became ancestors of peoples living on the western border and intermixed with Omotic speakers to form the Ari, Basketo, Dimi and Gimira-Maji groups of tribes. The languages spoken in Ethiopia in the third millennium B.C. are derived from a single stock (Hamito-Semitic or Afro-Asiatic) originating in the Eastern Sahara.
-

- Shinasha young women (North Omotic speakers). DNA analysis of Shinasha, Shekkacho, Wolayta and other North Omotic-speaking individuals has revealed that they are closely related to other Afro-Asiatic speakers in Northeast Africa. The anthropologist/linguist Harold C. Fleming, who coined the term “Omotic”, could already deduce this from their physical resemblance to their Cushitic and Ethiosemitic-speaking neighbors. This affinity suggests that the Shinasha and other North Omotic speakers directly descend from the original Omotic settlers of the Horn (Gopalan et al. (2019); López et al. (2021)).
-

- An Ari woman (South Omotic speaker). In contrast to the North Omotic-speaking groups, the Ari and other South Omotic-speaking individuals are both phenotypically and genetically distinct from other local Afro-Asiatic speakers. The South Omotic speakers are instead similar to Niger-Congo and Nilo-Saharan-speaking populations inhabiting the Great Lakes region. This suggests that the Ari and other South Omotic speakers are language shifters i.e., their forager ancestors originally spoke non-Afro-Asiatic tongues (Gopalan et al. (2019); Boattini et al. (2013)).

Dermatoglyphic affinities of Afro-Asiatic and Nilo-Saharan-speaking populations in Ethiopia. Fingerprint patterns, which are genetically inherited, indicate that the North Omotic-speaking Shinasha share ties with the other sampled Afro-Asiatic-speaking individuals (Cushitic-speaking Oromo and Ethiosemitic-speaking Amhara and Tigray) rather than with the Nilo-Saharan-speaking individuals (Berta). This again suggests that the Shinasha and other North Omotic speakers directly descend from the original Omotic settlers of the Horn (Yohannes and Bekele (2015)).

Genomic analysis of various populations in Africa and Europe. The North Omotic-speaking Shekkacho sample has a similar level of West Eurasian ancestries (Afro-Asiatic & Iberian-related components) as the other Afro-Asiatic-speaking populations and Nubian groups in Northeast Africa. Although bearing some Mota-related admixture like the Ethiosemitic-speaking samples (Amhara & Tigray), the Shekkacho primarily belong to the Afro-Asiatic ancestral component, which peaks among Cushitic-speaking Somalis. This element is analogous to Hodgson et al. (2014)‘s “Ethio-Somali”, an inferred ancestral component that the scientists demonstrated via a battery of tests to be essentially West Eurasian. The foregoing supports the view that the earliest Omotic settlers in the Horn region were, like the ancient Cushitic pastoralists in the Great Lakes area, transplants from North Africa. As also observed by López et al. (2021), the North Omotic-speaking Shekkacho, Shinasha and Wolayta thus represent pockets of Omotic speakers who retained much of their original Afro-Asiatic ancestry, whereas this ancestry was largely diluted in South Omotic-speaking areas through intermixture with and assimilation of local hunter-gatherers. Since Omotic is regarded by most linguists as the first diverging branch of the Afro-Asiatic language family, this analysis holds implications for the population affinities of the Proto-Afro-Asiatic speakers (Gopalan et al. 2019)).
Genetic differentiation
Besides morphology and language, the postulation that the Pre-Nilote forebears of the Ari (who are presumably represented by the Mota specimen) were of a different ancestral stock than the progenitors of the Afro-Asiatic-speaking Ethiopian groups is strongly supported by genetic studies on these contemporary populations.
For starters, researchers have observed a high frequency of the derived SLC24A5 allele (Ala111Thr or rs1426654) among various Cushitic- and Semitic-speaking Afro-Asiatic populations in Ethiopia. This mutation is closely associated with lighter skin pigmentation, and is believed to have originated in or near West Asia. Almost 60% of Ethiopian Jews and ethnic Somalis from Somalia carry the variant. et al. (2015) found a similarly high percentage of the allele among the North Omotic-speaking Wolayta. By contrast, only 12% or so of the South Omotic-speaking Ari ironworkers possess the Ala111Thr polymorphism. Since the Mota specimen does not harbor the mutation, this suggests that the ancestors of the Afro-Asiatic-speaking groups in the Horn were responsible for having introduced the allele into the Ari gene pool. This, in turn, implies that these early Afro-Asiatic settlers were of a lighter complexion than the Ari’s Mota-like forebears.

Lactase persistence (LP) frequencies among various Afro-Asiatic-speaking Ethiopian populations and adjacent groups (Jones et al. (2015)).
Further evidence that the Mota specimen was clearly not ancestral to the local Afro-Asiatic-speaking populations can be seen through comparative lactase persistence (LP) analysis. While Llorente et al. observed that Mota does not carry any lactose tolerance alleles, such LP mutations are found at high frequencies among the region’s main pastoral groups. To this end, Jones et al. (2015) note that over 60% of Beja, Afar and Borana Oromos are lactase persistent, carrying several different LP variants. In their large study of lactose tolerance in eastern Africa, Tishkoff et al. (2007) likewise find widespread lactase persistence among not only the Afro-Asiatic-speaking populations, but also among certain Nilo-Saharan and Niger-Congo pastoralist groups (like the Datog Nilotes) that are known to have absorbed some early Southern Cushites in the Great Lakes region.

Various analyses showing negligible-to-low autosomal DNA affinities between the Ari and adjacent Afro-Asiatic-speaking populations (Vandorp et al. (2015)).
Most tellingly, the Afro-Asiatic-speaking populations in the Horn appear to have generally different autosomal DNA (auDNA) signatures than the Ari. Hodgson et al. (2014) observed that ethnic Somalis, Afar, Amhara, Tigray and Oromos are defined by a West Eurasian-affiliated ancestral component, which they refer to as “Ethio-Somali”. On the other hand, the Ari are defined by a separate, Nilo-Saharan-affiliated ancestral component, which the researchers call “Ethiopic”. The Wolayta, despite possessing comparatively greater Mota-like admixture, were again found to be closer overall to the Cushitic- and Semitic-speaking populations than to the Ari. Dobon et al. (2015) further confirmed the existence of a defining West Eurasian ancestral element among the Afro-Asiatic speakers in Northeast Africa. Vandorp et al. (2015), in turn, reaffirmed the lack of close ties between the Afro-Asiatic-speaking populations and the neighboring Ari.

Ethnolinguistic distribution of maternal lineages in the Horn, Nile Valley, Sahara, Maghreb, Great Lakes and the Arabian peninsula (Boattini et al. (2013)).
Similarly, Boattini et al. (2013) observed that the mtDNA of the South Omotic-speaking Dawro-Konta and Hamer in southern Ethiopia, who are closely related to the Ari, is tied to that of Nilotic and Bantu populations in the Great Lakes region as well as the Ongota (who are believed to have once spoken a Nilo-Saharan language). On the other hand, the maternal lineages of the Cushitic- and Ethiosemitic-speaking groups of the Horn cluster instead with those of other Afro-Asiatic-speaking populations in the Nile Valley, Yemen and Sahara.
Altogether, this is consistent with the aforementioned tradition that the Ari’s “Shanqilla” ancestors (i.e. Mota) were originally Nilo-Saharan speakers, who at some point in antiquity adopted an Omotic language from early Afro-Asiatic-speaking settlers.
Afro-Asiatic ancestral population
In their study of the Mota remains, Llorente et al. note that contemporary Ethiopian populations (specifically, the Afro-Asiatic speakers) have substantial West Eurasian affinities that the Mota specimen does not appear to possess. They therefore conclude that that ancestry likely arrived after Mota’s lifetime, and originated from a Sardinian-like population most similar to the early Neolithic farmers who settled Europe (according to f3 statistical analysis).
In lieu of modern reference populations, the researchers ran an admixture test using the Mota specimen and a Neolithic Linear Pottery culture or Linearbandkeramik/LBK sample (represented by the Stuttgart specimen) as the baseline African and West Eurasian samples, respectively, against which the DNA of the Ari and that of the neighboring Afro-Asiatic-speaking populations was compared. This alone constitutes something of a breakthrough in biogeographical analysis, where disparate, admixed modern groups are instead frequently used as stand-ins for ancient populations (e.g. Yoruba and Utah denizens versus early Africans and West Eurasians). Nonetheless, there remain several problems with utilizing these particular Mota and Linearbandkeramik samples as the ancient proxy groups. Perhaps the most conspicuous issue is that, despite their antiquity, neither the Mota fossil nor the makers of the Neolithic LBK industry are representative of the ancestral Afro-Asiatic speakers in the Horn. We already examined above why that is vis-a-vis the Ari and their apparently Nilo-Saharan-speaking Mota progenitors, so let us now turn to the Linearbandkeramik agriculturalists.
While the LBK farmers may very well have prehistorically contributed some genes to the populations in Northeast Africa (and/or elsewhere on the continent), they do not appear to have been responsible for the majority of the West Eurasian affinities that are found today in the Horn. There are a number of reasons why we can be confident of this. Firstly, although the mtDNA haplogroup N1a, which is quite common today among the Afro-Asiatic-speaking populations in the Horn, is found at high frequencies among Neolithic LBK samples from Central Europe, the particular 7,000 year old Stuttgart sample from Germany that Llorente et al. utilized in their own analysis belongs to the T2 clade (cf. Lazaridis et al. (2013)). This latter maternal haplogroup is, by comparison, relatively rare in Northeast Africa. Unfortunately, since the Stuttgart sample is that of a woman and females do not inherit a Y chromosome, it is unclear whether the situation is the same with regard to the paternal side of things. Haak et al. (2010) did, however, successfully type three Neolithic LBK males for their Y-DNA. These individuals were assigned to haplogroups G2a3 and F*, both of which are also uncommon in Northeast Africa.
Another clue that the Linearbandkeramik agriculturalists are not the main source of the West Eurasian ancestry in the Horn is the fact that local rock art, such as at Laas Geel in northern Somalia, clearly depicts both herders and their domesticated cattle prior to the ~3,000 year old date that Llorente et al. propose as the arrival period of the LBK people. Those cave paintings also have stylistic precedents in the Arabian Peninsula (as explained further here, under ecology, rock art and genetics). Moreover, the Stuttgart woman, like the Mota man, was apparently unable to digest lactose. Lazaridis et al. remark that she lacks any alleles associated with lactase persistence. This in itself is not especially unusual for members of non-herding populations (whether farmers or hunter-gatherers) since they often do not drink cow milk and thus their bodies have no need to continue producing the lactase enzyme passed the breast-feeding infant age. Nevertheless, the finding is at odds with the aforediscussed high frequency of lactose tolerance among the pastoralist Afro-Asiatic-speaking groups and also contrary to the great diversity of the LP alleles present in the region.

The ~3,500 year old Cairn 4 burial excavated by Daniel Stiles, containing the 190 cm/6’4″ skeleton of an early Cushitic male settler (Stiles and Munro-Hay (1981)).
Aside from differing genetic markers and pastoral rock art, archaeology provides the most obvious indication that a West Eurasian-affiliated population — one likely distinct from the LBK farmers — co-existed in the region at around the same period as the Mota man and his kinfolk. In the 1980s, at various oases on the eastern margin of the Chalbi desert, the anthropologist and archaeologist Daniel Stiles excavated a number of cairns belonging to the Cushitic-associated Savanna Pastoral Neolithic (Stone Bowl Culture). The burials yielded the remains of tall individuals of “Caucasoid” physical type, with the oldest such skeleton, a 190 cm/6’4” male, radiocarbon-dated to around 3,500 years before present. By contrast, Llorente et al. estimate the roughly coeval Mota specimen’s stature at only 154.8 cm/5’1” i.e., a whopping 35.2 cm/13.9 inch difference in height! As explained in detail on The Elongated African fallacy (under physiognomy and exotic influences), indirect evidence has also established that, unlike Mota, these early Cushitic settlers possessed both the Ala111Thr allele for lighter skin pigmentation and a lactase persistence mutation as well as non-kinky (cymotrichous) hair texture.

Approximate difference in height between the Savanna Pastoral Neolithic Cairn 4 specimen and the coeval Mota hunter-gatherer specimen (MrInitialMan.com).
Thus, with the discovery, biological examination and dating of the Mota fossil, we now have concrete evidence of the existence of at least two genetically and physically distinct populations residing in the region during the Neolithic: one population ancestral to the Afro-Asiatic-speaking groups and linked with West Eurasians, and the other population ancestral to the Aroid groups and possibly linked with Nilo-Saharans (though the Mota specimen’s diminutive stature, his hunter-gatherer lifestyle, and his and the Ari’s genetic ties with the Sandawe may point to deeper Khoisan relations).
Unidentified East Eurasian elements
In addition to unrepresentative reference populations, another problematic aspect of the Mota biogeographical analysis is its intrinsic assumption that two proxy groups — an African sample and a West Eurasian sample — are sufficient to accurately capture the ethnic composition of the populations in Northeast Africa and elsewhere on the continent. A closer, multidisciplinary look at the broader data, however, points to the presence of at least a third ancestral element; one with apparently East Eurasian affinities.
Morphology and ancient testimonies

Craniometric relationships between various ancient and modern populations (Brace (1993)).
Almost two centuries’ worth of craniometric and anthropometric studies contradict the suggestion in the Mota paper that the Afro-Asiatic-speaking populations in the Horn may have been ancestrally formed through interbreeding between an early Sardinian-like population (the LBK makers) and a Mota-like population. In actuality, rather than morphologically clustering somewhere halfway between these two ancient reference groups — as one would expect for peoples that are supposedly 50% European and 50% African — the Afro-Asiatic speakers show few of the osteological characteristics of actual biracial populations. They instead appear “skeletally Mediterranean” (see, for instance, Brace (1993) to the right; also Kemp (2006) and the other studies here).

Sidamo men of relatively “pure” Cushitic type, closely resembling many Oromos.
-

- Hadiya Sidamo leader Gaaxi Ani Fit. Geejja Garbo with the mariin (“red-brown”) phenotype widespread among the Cushitic peoples.
-

- A Sidamo man of mixed Cushitic and South Omotic ancestry. The South Omotic or Ari-like forager influence is particularly evident in the kinky hair texture and broader/flatter nasal bones and cartilage, as well as what Carleton Coon refers to as “an enlargement of both sagittal and lateral diameters of the face.”
-

- Mengistu Haile Mariam, Ethiopia’s former Head of State, was born to Amhara and Shanqilla parents. His physiognomy similarly reflects this mixed Ethiosemitic-South Omotic parentage.
The anthropologist Carleton Coon observed this firsthand in his detailed examination of physical types in the Horn, “The Mediterranean Race in East Africa”, a chapter in his influential 1939 work The Races of Europe. By analyzing the Sidamos of southern Ethiopia in particular, who are the actual product of recent intermixture between Cushitic peoples and adjacent Pre-Nilotes, he was able to see what exact morphological changes such hybridization produces. Coon thus concluded that the local “Hamitic” populations were essentially “Caucasoid” and that “Negroid” influence, while also present, was on the whole minor.
Furthermore, Coon was able to identify an additional “non-Negroid” ancestral element, which was especially important among Somalis, Afars/Danakils, Agaus and other Cushitic groups. This swarthy “Veddoid” component, he postulated, may have arrived in antiquity from the Indus Valley by way of Southern Arabia along with the zebu cattle (Bos indicus):
Later than the development of highland agriculture in East Africa was the introduction and diffusion of pastoral nomadism. The cattle complex, with its elaborate set of social restrictions and of social differentiation on the basis of wealth in herds, was introduced from India by way of southern Arabia, along with the humped zebu, at some none too distant period, probably as late as the first millennium B.C.[…]
Our survey of the metrical characters of the inhabitants of the Hamitic racial area has brought several facts to light; the agricultural population of the Ethiopian highlands, both indigenous and imported from Arabia, belongs to a tall, dolichocephalic to mesocephalic, leptoprosopic, moderately leptorrhine race, which is Mediterranean in metrical position and cannot be distinguished, on the basis of the more commonly taken measurements, from blond and brunet Mediterraneans of Europe and North Africa. The Somalis, on the other hand, belong to an extreme racial form; extremely linear in bodily build, extremely narrow-headed and narrow-faced, with a special narrowness of the jaw. The relationship of the Somalis, on metrical grounds, is with some of the peoples of India as much as with the Mediterraneans elsewhere. The leptosome tendency, and the narrowness of the face, remind one of the same tendency found among the mixed Bedawin group of the Hadhramaut. It cannot be attributed to negro-white mixture, for that phenomenon, as witnessed among the Sidamos, has produced a heaping of characters, resulting in an enlargement of both sagittal and lateral diameters of the face, in some cases in excess of either the Hamitic white or the negroid parent. Upper face height and nose height are especially affected. The Somali face and nose are not long, they are merely narrow.

Royal effigy of the Meroitic King Asharramon (Fig. 191) and heads of ancient Egyptian commoners of similar “Austral-Egyptian” type (Morton (1854)).
Coon’s assertion is nothing new, for various early Greek, Roman, Egyptian and Brahmin writers, including Strabo and Philostratus, inform us of ancient colonies of Indus Valley peoples in Northeast Africa. For example, Bahadur (1917) notes that “Eusebius states that Ethiopians emigrating from the River Indus settled in the vicinity of Egypt [Meroë].” Nilus similarly relayed to Apollonius Tynaeus that “the Indi are the wisest of all mankind. The Ethiopians [Meroites] are a colony from them: and they inherit the wisdom of their forefathers”.
Herodotus (c. 440 BCE) indicates that different populations — both light-skinned and dark-skinned — lived in ancient Aethiopia, an area in Africa roughly corresponding with the territory to the south of Egypt and Libya. Among these inhabitants were (cf. Herodotus: The Histories):
- the “Ethiopians nearest to Egypt”, who were centered in the “great city called Meroe, which is said to be the capital of all Ethiopia” i.e., the ancient Meroites of Nubia (Book II: chapters 1‑98, Book III: chapters 89‑117); ancient DNA analysis indicates that the Meroites were closely related to the ancient Egyptians, with both populations carrying a predominant West Eurasian ancestry — see Ancient DNA from Sudan
- the “nomad Ethiopians”, who lived along a great lake near Elephantine island in Upper Egypt i.e., early Beja Cushites (Book II: chapters 1‑98)
- the “long-lived Ethiopians”, who “dwelt on the Libyan coast of the southern sea” and to whom the Persian king Cambyses sent envoys; they were “said to be the tallest and fairest of all men” and the “tallest and fairest and longest-lived of all men” i.e., the ancient Macrobians (Book III: chapters 1‑38, Book III: chapters 89‑117); ancient DNA analysis has confirmed that these early Cushitic settlers in East Africa were of North African origin, carrying a predominant West Eurasian ancestry (including a derived allele associated with lighter skin pigmentation) — see discussion below
- the “black-skinned” Ethiopians, who in complexion resembled the “Indians [that] dwell far away from the Persians southwards” (Book III: chapters 89‑117)
- the Ethiopians “who dwell about the holy Nysa”, located in the mountains of Upper Nubia (Book III: chapters 89‑117)
- the “Ethiopians from the east”, who lived in Asia (in or near Balochistan) and “are straight-haired” (Book VII: chapters 57‑137)
- the “Ethiopians of Libya”, who dwelled “in the south of Libya” and “have of all men the woolliest hair” i.e., Niger-Congo & Nilo-Saharan-speaking peoples/negroes (Book IV: chapters 145‑205, Book VII: chapters 57‑137)
- the “cave-dwelling Ethiopians”, whose “speech is like none other in the world[…] it is like the squeaking of bats” i.e., click-speaking Khoisan (Book IV: chapters 145‑205)
Other anthropologists have likewise proposed that an ethnographical tie exists between early Afro-Asiatic-speaking populations in Northeast Africa and the makers of the Indus Valley civilization. Following detailed examination of the skulls and murals of the ancient Meroites in the Nile Valley, Samuel George Morton asserted that “the Austral-Egyptian or Meroite communities were an Indo-Arabian stock, engrafted on the primitive Libyan inhabitants”. He further explained that, while such an influence was also present among commoners in Egypt, it was strongest in Nubia, including among the ruling class:
I observe, among the Egyptian crania, some which differ in nothing from the Hindoo type, either in respect to size or configuration. I have already, in my remarks upon the ear, mentioned a downward elongation of the upper jaw, which I have more frequently met with in Egyptian and Hindoo heads than in any other, although I have seen it occassionally in all the races. This feature is remarkable in two of the following five crania (A, B), and may be compared with a similar form from Abydos[…]
It is in that mixed family of nations which I have called the Austral-Egyptian that we should expect to meet with the strongest evidence of Hindoo lineage; and here, again, we can only institute adequate comparisons by reference to the works of Champollion and Rosellini. I observe the Hindoo style of features in several of the royal effigies; and in none more decidedly than in the head of Asharramon (Fig. 191), as sculptured in the temple of Debod, in Nubia. The date of this king has not yet been ascertained; but, as he ruled over Meroe, and not in Egypt, (probably in Ptolemaic times [B. C. 200-300],) he may be regarded as an illustration of at least one modification of the Austral-Egyptian type[…]
Another set of features, but little different, however, from the preceding, is seen among the middling class of Egyptians as pictured on the monuments, and these I also refer to the Hindoo type. Take, for example, the four annexed outlines (Fig. 192), copied from a sculptured fragment preserved in the museum of Turin. These effigies may be said to be essentially Egyptian; but do they not forcibly remind us of the Hindoo?
The Badarian factor

A typical Badarian male skull (Stoessiger (1927)).
Of all the ancient populations in Northeast Africa, the Badarians of Upper Egypt were most often regarded as the likeliest link between the local Afro-Asiatic-speaking groups and the Indus Valley peoples.
The Badarian culture flourished between 4400-4000 BCE. It consisted of small villages of semi-nomadic agropastoralists, who kept domesticated animals, cultivated grain, produced the first glazed objects, and also used metal. The Badarians interred their dead in cemeteries on the outskirts of their living area. Within the actual graves, the deceased were laid out in the fetal position, facing the setting sun toward the west. They were buried with some clothing items, pottery, jewelry and a fertility idol. Despite its brief existence, the culture is considered archaeologically important since Badarian sites have yielded the earliest evidence of agriculture in Egypt.
In Coon’s examination of Badarian skeletal remains, he observed that they were on the whole quite similar to those of other predynastic Egyptians. He described the Badarians as essentially “Mediterranean” in the anthropological sense, possessing cranial affinities with both the Afro-Asiatic-speaking populations of the Horn (although the Horn groups are ultimately closer to the later Naqadans) and the Dravidian populations of southern India:
From the type site, Badari, come the earliest skulls of a definitely Egyptian group which have yet been discovered. These Badarians lived about 4000 B.C., after the climate had become considerably drier than it was in Tasian times, so dry, in fact, that in many cases the skin and hair of their dead have been naturally preserved. The skin was apparently brunet white, while the hair was black or dark brown in color, thick, of fine texture, and usually wavy in form.
Although the Badarians, like the Tasians and Merimdians, still hunted and fished to enhance their larders and vary their diet, they lived primarily by agriculture and by herding cattle and sheep. Unlike the Merimdians they raised no pigs. By hammering copper they were entering the transition from the Neolithic to the Metal Age. They navigated the Nile in ships, whose shapes are revealed by pottery models, but we cannot be sure that they sailed them. These Badarians were undoubtedly newcomers to Upper Egypt who displaced the Tasians and perhaps other predecessors.[…]
The Badarian type represents a small branch of the Mediterranean racial group. The head is unusually high in comparison to the other dimensions, and the facial skeleton is in the absolute scale unusually small; the mandible is small, narrow and light. Its mean male bicondylar diameter is the smallest known, while the bigonial diameter of 91.6 mm. is also extremely low.[…]
Morant shows that the Badarian cranial type is closely similar to that of some of the modern Christians of northern Ethiopia, who incidentally do not show negroid characteristics in the skull, and also to the crania of Dravidian-speaking peoples of southern India. One might add that living Somalis show a close approximation to this physical type in most respects, and the extremely narrow jaw in which the Badarians seem to reach a world extreme may be duplicated among both Somalis and the inhabitants of southern India. In Europe, the closest parallel to the Badarian type is found among modern Sardinians, but this is not as close as their relationships to outer and later Egyptians.
Coon indicates that the Badarians seem to have eventually been absorbed by the ensuing predynastic Egyptians of Naqada. He also asserts that a Badarian strain persists in the Horn. Despite the prevailing “Mediterranean” element, this lingering influence, he suggests, manifests itself in various ways; particularly through attenuation of the distal or remote segments of the limbs (i.e., small wrists, hands, ankles and feet), as is common among the populations of southern India:
The bodily build of the African Hamites is typically Mediterranean in the ratio of arms, legs, and trunk, but the special attenuation of the extremities among the Somalis is a strong local feature, which finds its closest parallels outside the white racial group, in southern India and in Australia.
In her comprehensive study of Badarian skeletal remains, the anthropologist Brenda Stoessiger (1927) confirmed that the Badarians were closely related with other predynastic Egyptian series, but also bore relations with Dravidian populations in the Indian peninsula. She attributes these bonds to parallel population movements, westward into Northeast Africa and eastward into South Asia, from a common center in or near the ancient Caucasus:
Anthropologists have frequently drawn attention to the similarity in the appearance of the skulls of the Hindu races of India and the Early Egyptians. In a paper “Sur l’Origine de l’Ancienne Race Egyptienne,” published in the first volume of the Mémoires de la Société d’Anthropologie de Paris, pp. 410-422, Pruner-Bey notes this similarity only to reject it; each race, he says, has a skull, small and oval in shape; the body and extremities are for both races beautifully proportioned; but there is a marked difference in the fleshy parts, the Ancient Egyptian resembling the modern Berber, while the Hindu is bronze to bistre in colour. Finally he dismisses the idea of direct relationship on the grounds of linguistic differences, an argument which would scarcely now-a-days be advanced.[…]
From skull measurements alone it would be difficult to choose between the primitive Indian and Egyptian series as the group to which the Badarians are closer. Unfortunately it is not possible to carry the analogy further and find low coefficients between the later Egyptian series and the Indian series as the types diverge in different directions.[…]
This study confirms the conclusions based on cultural and topographical evidence that the Badarian skulls are early Predynastic Egyptian but if anything more primitive in type than the other series of this period, though the mean direct measurements differ very little from them. The early and late Predynastic types however do show a significant difference which, if we may assume the races to be divided by a period of four or five thousand years or more, may be accounted for by slight evolutionary changes or a gradual infusion of races.
How far do these results confirm Sir Flinders Petrie’s theory of a Caucasian origin? When we compare the Badarian race with others outside Egypt, it is not the Mediterranean or any Negro type which it resembles most closely but the primitive Indian, the Dravidian and the Veddah. Thus they do not oppose the suggestion of a common origin in the Caucasus from a race sending one branch westward to Egypt and Europe and another south-eastward to India. To confirm this, however, we should need series of ancient skulls from Palestine, Persia, and Western India.
Genetic and phenotypic oddities
According to Coon (1939), ethnic Somalis (over 86% among northern Darod individuals), Afars and other lowland Cushitic groups possess significantly higher rates of non-kinky hair texture than do the Amhara (40%), Tigray and other highland Abyssinian Semitic speakers. He asserts that the Sidamo, despite their elevated South Omotic admixture, also have a higher rate of non-kinky hair texture than do the Abyssinians. Puccioni (1931) reported comparable findings for the Somali clans in Somalia. Charpin and Georget (1977) similarly observed that only 3.8%-4.9% of the Issa Somalis and the Afars of Djibouti had hair in the kinky/ulotrichous class (cheveux crépus).
-

- Hair form and anthropometric means of the Afars and Issa Somalis of Djibouti. Just 3.8%-4.9% of these individuals have hair in the kinky/ulotrichous class (cheveux crépus) (Charpin and Georget (1977)). This is consistent with anthropological and archaeogenetic analyses, which have found that the Cushites of the Pastoral Neolithic, who bore a predominant Eurasian ancestry, had soft-textured hair (For details, see Ancient DNA from Sudan).
-
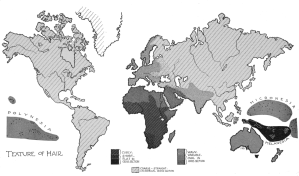
- Global hair texture distribution map. Many Afro-Asiatic-speaking individuals of the Horn of Africa (particularly Cushitic speakers in the northern areas) have wavy hair like other Afro-Asiatic speakers in North Africa and the Middle East.
Besides the osteological work, ancient testimonials, zebu cattle complex and hair morphology, there are a couple of other indications that an East Eurasian ancestral component likely exists among the Afro-Asiatic-speaking groups in Northeast Africa. Narasimhan et al. (2019) report a high prevalence of the Afro-Asiatic-linked E1b1b paternal haplogroup among Late Bronze Age/Early Iron Age individuals from the Swat Valley in northern Pakistan, specimens which also exclusively bore M and N mtDNA derivatives. This points to ancient ties between this area, Mesopotamia, the Levant, the Arabian peninsula, North Africa and the Horn of Africa (cf. Table S1-S5). More specifically, the presence of haplogroup E1b1b in South/Central Asia this early in time likely reflects eastward incursions by Neolithic and/or Bronze Age Levantines (the original Elamo-Dravidian settlers, perhaps?). These newcomers possibly interbred with local populations, before eventually returning to the Afro-Asiatic fold along with their newly-acquired East Eurasian genes. Such early connections between Afro-Asiatic-speaking populations and groups bearing East Eurasian ancestry are likewise supported by Witas et al. (2013), who analyzed Early Bronze Age to Roman era specimens excavated at Tell Ashara (Terqa) and Tell Masaikh (Kar-Assurnasirpal) in Mesopotamia. The researchers found that these ancient individuals (dated to 2500BCE-500CE) belonged to the mtDNA clades M4b1, M49 and/or M61, maternal lineages that are today concentrated and believed to have evolved in the Indian subcontinent. Various autosomal DNA analyses have also observed an East Eurasian component in their Somali samples (e.g. Kidd et al. (2011); Kidd (2011b); Truelsen et al. (2017); Pereira et al. (2017)). Other factors supporting this association are the elevated frequency of the B blood group among Ethiopian Somali males (44.44%) and Afar males (33.33%) (cf. Abegaz (2021)) as well as among Beni Amer Beja (31%; Corkill (1949)) — a serological system that is also common among Egyptians (24.1%; Swelem et al. (2018)) and Riverain & Shaigiya Sudanese “Arabs” (25% and 24%, respectively; cf. Corkill (1949)) and which globally peaks among populations in South Asia (Kocak et al. (2017)); the high mean number of private alleles that are found today in certain Cushitic groups (cf. Babiker et al. (2011)); as well as the presence in these populations of unusual genetic variants of apparent East Eurasian origin (e.g. the EDAR gene’s derived allele, which is associated with hair thickness among populations in eastern Asia; around 12.5% of ethnic Somalis bear the mutation, whereas it is largely absent in Subequatorial Africa, the Maghreb, West Asia and Europe).
(*N.B. Our genome analysis has also detected an East Eurasian component borne by our Cushitic, Ethiosemitic and North Omotic-speaking samples from the Horn. For specifics on this East Asia-associated element, see Genetic affinities of the Cushitic, Ethiosemitic and North Omotic-speaking populations of the Horn of Africa.)
-

- Autosomal SNP analysis detecting the presence of a notable South/Central Asian component (blue) in the Somali sample in contrast to the Ethiopian Jew and Mozabite samples (Kidd et al. (2011)). (*N.B. Kidd (2011b) indicates that this Somali sample is from the Yale collection, which ALFRED notes was originally gathered by Dr. Qasim Mehdi. This is the same cohort that Pagani et al. (2012) used in their analysis, and thus likely mainly consists of Hawiye Somali rather than Benadiri Somali individuals.)
-

- Autosomal SNP analysis again detecting a South/Central Asian component (purple) in the Somali sample. Oddly, this component is found at lower frequencies in the Iranian, Turkish and Druze samples, although these populations border Central Asia (Truelsen et al. (2017)).
-

- Autosomal SNP analysis yet again detecting a South/Central Asian component (orange) in the Somali sample. This component is also found in the Libyan sample, albeit at lower frequencies (Pereira et al. (2017)).

Genome analysis of modern Emirati individuals detecting the presence of a South/Central Asian component. Researchers have usually ascribed this element among contemporary Arabians to recent admixture with persons from South/Central Asia. However, the absence of the Ancestral South Indian component among Afro-Asiatic speakers in general, coupled with the presence of a South/Central Asian component among some Afro-Asiatic-speaking individuals, suggests deeper associations, perhaps linked to the dispersal of the original Elamo-Dravidian speakers (Elbait et al. (2021), Figure S2).

Ancestral composition of modern populations in South and Central Asia. The Onge or Ancestral South Indian genome component is today widely distributed across this region. However, it has not been observed among the Afro-Asiatic-speaking populations in Africa. This suggests that the South/Central Asian inferred element, which in some analyses has been detected among modern Cushitic, Berber and Arabic speakers, was either a) derived from an area in South/Central Asia, such as Balochistan, where the Ancestral South Indian component is not found at appreciable frequencies, or b) introduced early on by South/Central Asian settlers, such as the makers of the Indus Valley civilization, at a time period when the latter had not yet interbred with the original bearers of the Ancestral South Indian component (Lazaridis et al. (2016)). (*N.B. Distance analysis on the Vahaduo Admixture JS program indicates that, besides the recent IND_Great_Andamanese_100BP sample on Eurogenes’ official Global25 datasheet, the Onge share the nearest genetic affinity with the LAO_Hoabinhian specimen (see here). Ergo, the latter ancient individual is currently the best available stand-in for “pure” Ancestral South Indian ancestry.)
Final observations and recommendations
Another interesting facet of the Mota analysis concerns Llorente et al.’s assertion that they were able to detect West Eurasian affinities in every African population that they examined. While such ancestry is not unexpected for certain groups like the Maasai, Sandawe, and Khwe and Nama Khoi (who are known to have assimilated some early Cushitic pastoralists), it is less clear how isolated hunter-gatherer populations like the Xun/!Kung San and Mbuti Pygmies would have acquired such an influence. It is tempting to suggest that the spread of haplogroup E may have had something to do with this. However, the fact that the Neolithic LBK farmers apparently did not carry the paternal clade rules them out as the disseminating ancient population. So does the fact that the Stuttgart LBK woman possesses the derived SLC24A5 allele for lighter skin pigmentation, whereas most African populations, other than the Afro-Asiatic-speaking groups in the Horn and North Africa, do not.
Given the foregoing, better potential reference populations for future ancient DNA tests would be:
- For the source of the West Eurasian ancestry in the Horn – The oldest skeletons in the Savanna Pastoral Neolithic mound cairns that were excavated by Daniel Stiles. These remains belong to early Southern Cushitic settlers. The A-Group, C-Group and Kerma peoples of ancient Nubia and the predynastic Egyptians of Naqada are also interesting possibilities, as they are craniometrically quite close to present-day Afro-Asiatic speakers in the Horn. A tertiary option is the medieval-period KulR17 infant from Kulubnarti in Sudan, which Sirak et al. (2015) found to possess Middle Eastern affinities.
- For the source of the unidentified East Eurasian ancestry in the Horn – The ancient Badarians of the Nile Valley. Osteological analysis by Stoessiger (1927) suggests that they are as closely related to other predynastic Egyptian groups as they are to modern Dravidian populations in South Asia.
- For the source of the putative West Eurasian ancestry in Subequatorial Africa – The makers of the Iberomaurusian industry of Northwest Africa. They lived before the derived mutations for lighter skin pigmentation evolved, and also appear to have been lactose intolerant. As such, the Iberomaurusians easily could have spread West Eurasian genes to the similarly dark-skinned, lactose intolerant early hunter-gatherer populations in the adjacent subequatorial areas. The makers of the Capsian culture are also an intriguing alternative for comparable reasons, as are the Kiffians and Tenerians of the Gobero in the Sahara (who are presently being typed for ancient DNA, according to paleontologist Paul Sereno).
For related archaeogenetics, see Ancient DNA from Sudan. Also stay tuned for new aDNA work from Egypt.
*Update #1
In January 2016, Llorente et al. published an erratum online pertaining to their Mota study. The announcement (summarized here by Scientific American) indicates that there was a software-related oversight on the researchers’ end, which caused them to unknowingly overlook some chromosomal affinities that do apparently exist between the Mota specimen and their West Eurasian reference sample. The scientists also write that the proposed large ancient migration from West Eurasia was instead mainly confined to East Africa:
Erratum to Gallego Llorente et al. 2015
The results presented in the Report “Ancient Ethiopian genome reveals extensive Eurasian admixture throughout the African continent“ were affected by a bioinformatics error. A script necessary to convert the input produced by samtools v0.1.19 to be compatible with PLINK was not run when merging the ancient genome, Mota, with the contemporary populations SNP panel, leading to homozygote positions to the human reference genome being dropped as missing data (the analysis of admixture with Neanderthals and Denisovans was not affected). When those positions were included, 255,922 SNP out of 256,540 from the contemporary reference panel could be called in Mota. The conclusion of a large migration into East Africa from Western Eurasia, and more precisely from a source genetically close to the early Neolithic farmers, is not affected. However, the geographic extent of the genetic impact of this migration was overestimated: the Western Eurasian backflow mostly affected East Africa and only a few Sub-Saharan populations; the Yoruba and Mbuti do not show higher levels of Western Eurasian ancestry compared to Mota.
We thank Pontus Skoglund and David Reich for letting us know about this problem.
First off, the researchers are to be commended for their professionalism; both for having the courtesy to admit to a mistake (which happens) and then promptly trying to redress that, and for having the vision to conduct such an ancient DNA analysis in the first place.
That said, what are the implications of the Mota erratum? Not much since, as explained above, Mota was not an adequate African proxy to begin with. His uniparental lineages already pointed to gene flow from an early Afro-Asiatic-speaking settler group(s), as is the situation with his contemporary Ari relatives. This introgression is now also supported by Llorente et al.’s revised analysis, which has apparently detected minor West Eurasian ancestry in Mota’s genome.
A better and older African reference population than Mota would, therefore, perhaps be Schepartz (1987)’s hunter-gatherer sample from the Gogoshiis Qabe Rockshelter in southern Somalia (8,100-5,400 BP). Sellers (2008) compared these skeletons’ dental metric affinities to those of modern ethnic Somali pastoralists and Final Paleolithic Nubian hunter-gatherers, Sudanese agriculturists (3,400-1,200 BP) and Sudanese intensive agriculturists (1-1,500 BCE), and found that the ethnic Somali individuals were much more closely related to the more recent Sudanese intensive agriculturalists (viz. the Meroitic, X-Group and Christian period populations) and Sudanese agriculturalists (viz. the A-Group and C-Group populations) than to the Gogoshiis Qabe Rockshelter hunter-gatherers and Final Paleolithic Nubian hunter-gatherers. As explained on Ancient DNA from Sudan, this is because ethnic Somalis and other Afro-Asiatic-speaking populations that today inhabit the Horn did not actually evolve there. Their ancestors — ancient Cushitic speakers; not the Gogoshiis Qabe Rockshelter hunter-gatherers or Mota or the LBK makers — arrived instead from the Sahara and Nile Valley. The latter area is the likely Afro-Asiatic urheimat or “original homeland”.
For a comprehensive overview of these early “Hamitic” settlers in the Horn, see Punt: an ancient civilization rediscovered.
*Update #2
It is 2020, a good four years since our last update, and much has happened in the world of palaeogenetics. We now have genomic data from both the Iberomaurusians in Northwest Africa and early Southern Cushitic settlers in East Africa, as well as more extensive testing of ancient Egyptian specimens of the Nile Valley. These ancient DNA analyses, amplified by studies on modern groups, have gone a long way towards helping to clarify existing and past population relationships and inheritance patterns.
Anatolian Neolithic component & ancestral deconvolution
As discussed above, various SNP-based autosomal DNA analyses have suggested that the modern Afro-Asiatic speakers in the Horn of Africa (and to a lesser extent the Afro-Asiatic speakers in North Africa) harbor two distinct ancestries: one non-African and the other African.
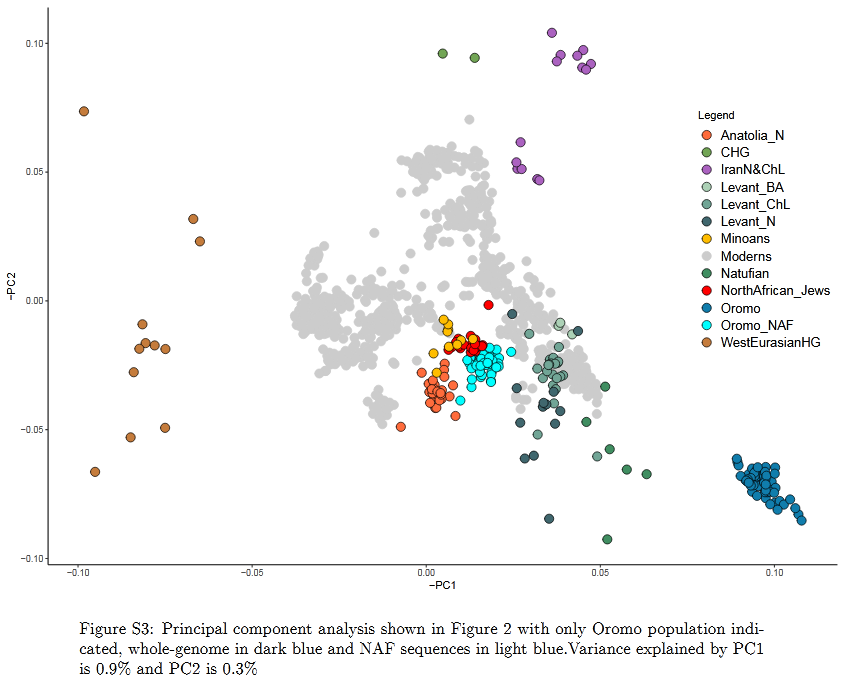
Non-African ancestral composition of the Oromo. This NAF component is estimated to be comprised of 85% Anatolian Neolithic and 15% CHG for the Oromo, Amhara and Wolayta, and 92% Anatolian Neolithic and 8% CHG for the Somali (Molinaro et al. (2019), Supplementary Information).
Researchers have hypothesized that this non-African component is associated with either Neolithic Europe (as represented by the LBK culture), the Mesolithic Levant (Natufian culture), or the Neolithic Levant (Pre-Pottery Neolithic culture). According to Molinaro et al. (2019), when the whole genome of the Cushitic and Semitic-speaking individuals is analyzed (that is, when both the non-African and African components are examined together), their West Eurasian ancestry appears to be primarily derived from the Neolithic Levant. However, when the non-African component (NAF) is isolated through a process of ancestry deconvolution, it instead appears to be largely composed of the Anatolian Neolithic component, with minor admixture from the Caucasus Hunter-Gatherer (CHG) component. Among populations in Ethiopia, Molinaro et al. estimate this non-African component ratio at 85% Anatolian Neolithic and 15% CHG for the Amhara, Oromo and Wolayta, and 92% Anatolian Neolithic and 8% CHG for the Somali.
What are the merits of this biogenesis scenario?
- Molinaro et al. (2019) is the first study to attempt isolating the non-African component from the African component. As the scientists point out, this is a necessary analytical step because in whole genome analysis, the African component (due to its highly divergent nature) skews the overall allele frequency spectrum. Molinaro et al.’s thesis is strengthened by the fact that it has been successfully replicated by Aneli et al. (2021), the second and only other study so far that has applied ancestral deconvolution to the question. Like Molinaro et al., Aneli et al. found that the distal source of the West Eurasian ancestry borne by the Horn’s Afro-Asiatic-speaking populations is largely Anatolian Neolithic-related. The researchers call this ancestral signature “Pan-Mediterranean” since they found it to be characteristic of earlier groups throughout the Mediterranean, including the ancient Daunians of Apulia and the Minoans of Crete (*N.B. Recent studies have found pre-Ptolemaic Egyptian samples to be the best-fitting surrogate for the proximal source of the Horn groups’ West Eurasian ancestry (cf. Almarri et al. (2021), Table S4; Sirak et al. (2021)). This, in turn, implies that Anatolian Neolithic ancestry would have been abundant in Egypt too prior to the spread of haplogroup J carriers into the Nile Valley.)
- Among modern populations, the researchers identify Tunisian Jews as the closest in terms of ancestral composition to the non-African component borne by the Afro-Asiatic speakers in the Horn of Africa. This is supported by Serra-Vidal et al. (2019), which, like Molinaro and colleagues, found the Anatolian Neolithic component to be the defining non-African component among the Afro-Asiatic-speaking populations of the Maghreb.
- Among ancient populations, the researchers identify the Minoans of Crete as the closest in terms of ancestral composition to the non-African component borne by the Afro-Asiatic speakers in the Horn. This is supported by ancient Egyptian iconography such as the Grand Procession mural at the tomb of Rekhmire, which, on adjacent panels, depicts quite similar-looking ancient Puntite and Cretan figures (see Punt: an ancient civilization rediscovered).

Meshwesh, one of the ancient tribes of Sea Peoples. Note the braided hairstyle, which is still maintained by some Afro-Asiatic speakers in Northeast Africa (Salimbeti).
- It explains why in whole genome statistical modeling Levantine Neolithic samples are often a better proxy for the non-African ancestry present in the Horn region than are Mesolithic Natufian samples (e.g. Skoglund et al. (2017)). According to Schuenemann et al. (2017), the Levantine Neolithic component is itself composed of Natufian ancestry and later-arriving Anatolian Neolithic admixture. Hence, the Levantine Neolithic signal that is being picked up could in reality be associated with that component’s Anatolian Neolithic element rather than its Natufian element. This appears to be why, after ancestral deconvolution, Molinaro et al. primarily detected an Anatolian Neolithic affinity for the Horn’s West Eurasian ancestry but no Natufian affinity.

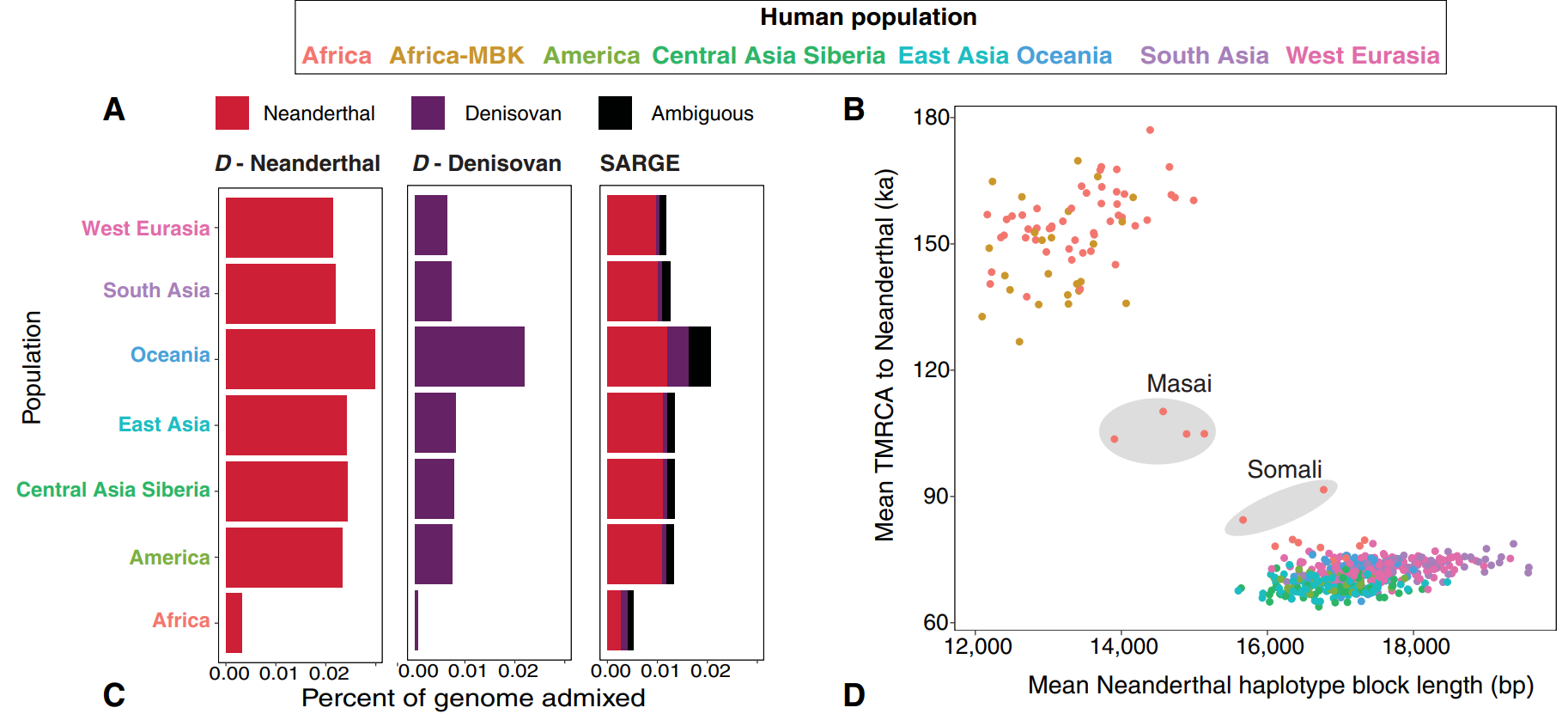
- It is supported by data on Neanderthal admixture. Schaefer et al. (2021) compared Neanderthal genomes to those of various modern populations, identifying haplotype block lengths that are indicative of Neanderthal genetic introgression. The scientists observed that their Somali, Mozabite Berber and Sahrawi samples were distinct from their other African samples and instead had haplotype block lengths similar to their non-African samples. This indicates that the early Cushitic settlers in the Horn region descended from a non-African population, though not the Natufians since the Natufians have been found to be devoid of Neanderthal admixture. More specifically, Schaefer et al. suggest that “these Neanderthal haplotype blocks may have originated in ancient European migrants to eastern Africa,” in agreement with the view that Minoan-related settlers (viz. the early Cushites) introduced the Anatolian Neolithic component to the Horn.

Mean Neanderthal haplotype block lengths, which identify Neanderthal admixture in various modern populations. The Somali sample has haplotype block lengths similar to those of the North African (Mozabite Berber and Sahrawi) and non-African samples. Schaefer et al. (2021) suggest that this is because “these Neanderthal haplotype blocks may have originated in ancient European migrants to eastern Africa.” This is consistent with the view that the early Cushitic settlers in the Horn of Africa descended from a population bearing Anatolian Neolithic ancestry (Schaefer et al. (2021)).
- It is supported by serological data on Rh-negative blood. Rhesus negative persons are nowadays quite rare, with around 89%-95% of examined individuals worldwide being Rh-positive. Rhesus negative blood has been observed at highest rates among European populations (~17% in Britain), especially Basques (29%). Equally high Rh-negative frequencies have also been reported among Berbers in the High Atlas mountains of Morocco (29%; cf. Weinstock (2014)), as well as in Syria (30.5%; Kocak et al. (2017)) and Al-Jouf province in Saudi Arabia (29%; Eweidah et al. (2017)). Golassa et al. (2017) likewise observed a high frequency of Rh-negative blood (21.32%) among the Afro-Asiatic-speaking residents of Gambella town, a Nilotic majority area located in the Gambella woreda (district) of southwestern Ethiopia (cf. Table 1). Of these Afro-Asiatic-speaking inhabitants (locally known as degegna or ‘highlanders’), most are Cushitic Oromo individuals (20.13%) (MANR (2016)). Similarly, Abegaz (2021) found elevated Rh-negative frequencies among his Oromo (34.74%) and Tigray (28.57%) female samples collected in Ethiopia’s northern Amhara region. Since Basques and Berbers have been found to be defined by the Anatolian Neolithic ancestral component (cf. Sarno et al. (2017), Supplementary Fig. S3; Serra-Vidal et al. (2019)), these findings accord well with Molinaro et al.’s thesis that Minoan-related settlers introduced the Anatolian Neolithic component to the Horn.


Lactase persistence allele frequencies of Afro-Asiatic-speaking populations in the Nile Valley and East Africa. The Northeast African-affiliated G-13907 variant peaks among the Beni Amer Beja (25%), the Arabian-affiliated G-13915 variant climaxes among Somalis (50%), and the South Cushitic-affiliated C-14010 variant is prevalent among the Yaaku (53.6%) (Tishkoff et al. (2010)).
It is supported by the lactase persistence (LP) alleles that are presently distributed in Northeast Africa. Tishkoff et al. (2010) and Hassan et al. (2016) note that the local Afro-Asiatic-speaking populations have high frequencies of various LP mutations, especially the pastoralist Cushitic groups. Of these derived alleles, the most commonly borne ones are: G-13907, which is centered in the Nile Valley and Horn of Africa and associated with cattle pastoralism; G-13915, which is centered in Arabia and associated with camel pastoralism; and C-14010, which is centered in the Great Lakes region and also associated with cattle pastoralism. The first two of these lactose tolerance variants are prevalent among North Cushitic and East Cushitic groups (G-13907 climaxes among Beni Amer Beja at 25%, and G-13915 peaks among Somalis at 50%), whereas C-14010 is typical of Southern Cushitic groups (the earliest occurence of this allele is among ancient Cushitic specimens of the Pastoral Neolithic; cf. Prendergast et al. (2018)). A fourth LP allele, T-13910, is most common in Europe and is also found among Tuareg Berbers, Mozabite Berbers and Fulani groups. According to Enattah et al. (2008):
The European T-13910 and the earlier identified East African G-13907 LP allele share the same ancestral background and most likely the same history, probably related to the same cattle domestication event.
Kulichová et al. (2017) indicate that the T-13910 LP allele was likely spread by pastoralists originating from outside Africa. Furthermore, Marcus et al. (2020) suggest that such population movement(s) from Europe into Africa also introduced both the R1b-V88 paternal haplogroup and Sardinian-related ancestry (Anatolian Neolithic). Given the shared ancestral background of the T-13910 and G-13907 lactase persistence variants, this implies that the Cushitic groups likewise inherited the G-13907 allele from ancestors bearing the Anatolian Neolithic component.

Haplotype network of lactase peristence (LP) alleles in Africa, Europe, the Middle East and Central Asia. The Northeast African G-13907 variant (located on haplotype 21) and the European T-13910 variant (located on haplotype 24) are associated with an A haplotype background (Liebert et al. (2017)). According to Enattah et al. (2008), the fact that these LP alleles share the same ancestral background denotes that they have “most likely the same history, probably related to the same cattle domestication event.”


- It accounts for European mitochondrial haplotypes that have been observed in the Horn. For example, Comas et al. (1999) note that, among the Somali individuals they examined, “high frequency of A at position 73 (probe A1) has been suggested to be an exclusively European marker” and that “the presence of CRS mtDNA haplotypes (with different lengths of Cs at position 309) are typical European mtDNA haplotypes.” CRS here denotes the Cambridge Reference Sequence, which belongs to the mitochondrial haplogroup H. This mtDNA clade is today the single most common maternal lineage carried by Europeans. Biogenesis scenarios centered on Arabian or Levantine populations (including European-admixed groups) would not be able to adequately explain the presence of these European haplotypes in the Horn given that: 1) the Horn region’s defining non-African ancestry, after ancestral deconvolution, is apparently Anatolian Neolithic rather than Levantine Neolithic, and 2) Mathieson et al. (2015) observed that “the Anatolian Neolithic samples do not resemble any present-day Near Eastern populations but are shifted towards Europe, clustering with Neolithic European farmers (EEF) from Germany, Hungary, and Spain.” Those European haplotypes are thus more likely to have been brought by Anatolian Neolithic-defined European groups, who settled in the Middle East and from there resettled in Africa. Molinaro et al. propose that the Minoan-related Sea Peoples did just that.
- It is compatible with Hodgson et al. (2014), which is the first admixture analysis on the Horn of Africa populations to exclude SNPs in strong linkage disequilibrium. As Gandini et al. (2016) explain:
Hodgson et al. argue cogently that genome-wide dating methods based on linkage disequilibrium are strongly biased in favour of recent admixture events.
Such recent admixture events did affect the current Ethiosemitic speakers, when they shifted from speaking their original Cushitic Agaw languages and adopted South Semitic languages. In the process, they acquired the paternal haplogroup J through intermarriage with Sabaeans (33% among the Amhara). They also have haplogroup A from admixture with South Omotic/hunter-gatherer populations: 17%-20% of Amhara, up to 36% of Tigrinya, and as high as 41% of Ethiopian Jews (cf. Wood et al. (2005), Appendix A; Scozzari et al. (2014), Table S7; Cruciani et al. (2002), Table 2; Gebremeskel (2018), Table 3.7).

Y-chromosome haplotype frequencies in select African populations Cruciani et al. (2002), Table 2). Most of the Ethiopian Jew individuals (41%) bear the haplotype I, which corresponds with the paternal haplogroup A. In Northeast Africa, this archaic African lineage is most common among Omotic and Nilo-Saharan-speaking populations. This suggests that the ancestors of the modern Ethiosemitic-speaking groups in Ethiopia significantly admixed with neighboring hunter-gatherers. Later periods of interbreeding with haplogroup J carriers from Arabia would dilute that elevated forager admixture, in the process raising the Ethiosemitic speakers’ overall West Eurasian ancestry to a level similar to that of their Cushitic-speaking neighbors (cf. López et al. (2021); their Cushitic-speaking Agaw sample has comparable West Eurasian ancestry as their Ethiosemitic-speaking samples despite the fact that the Agaw have sustained less recent gene flow from the Arabian peninsula).

Inferred ancestral components among the Afro-Asiatic-speaking groups of the Horn of Africa and other global populations. After correcting for linkage disequilibrium bias, most of the Cushitic and Ethiosemitic-speaking populations have non-African ancestry estimated in the 60%-70% range, similar to southern Moroccans. This non-African ancestry in the Horn largely belongs to the “Ethio-Somali” component, a West Eurasian ancestral element that peaks among ethnic Somalis (Hodgson et al. (2014), Table S6). (*N.B. The Sudanese sample above consists of Nilotes. It does not include Nubians or Sudanese “Arabs”, who instead share affinities with the Afro-Asiatic-speaking populations in Northeast Africa (see Dobon et al. (2015)).)

Pairwise FST estimates show the non-African affinities of the Ethio-Somali ancestral component. The Ethio-Somali component has the lowest pairwise FST values with the Maghrebi and Arabian ancestral populations (0.074, 0.083). FST values are on average lower between the Ethio-Somali component and non-African populations (mean=0.097) than with sub-Saharan African populations (mean=0.126) (Hodgson et al. (2014), Supplementary Text S1).
After correcting for this linkage disequilibrium bias, Hodgson et al. found that the non-African ancestry proportions for most of the Afro-Asiatic-speaking samples (Cushitic and Ethiosemitic speakers alike) are in the same 60%-70% range as southern Moroccans (Eaaswarkhanth et al. (2020) also observed a similar level of West Eurasian ancestry among many of the Kuwaiti Arab individuals they analyzed). Most of this estimated non-African ancestry in the Horn belongs to the “Ethio-Somali” component, a West Eurasian ancestral element that peaks among ethnic Somalis. The Ethio-Somali component correlates with ancient Cushitic ancestry, and is most closely related to the Maghrebi and Arabian ancestral components. Conversely, the African ancestry in the Horn primarily belongs to the “Ethiopic” component (also known as the “Omotic” component). This ancestral element correlates with indigenous hunter-gatherer or Mota-like ancestry, and peaks among the Ari Blacksmiths. The Ari today speak an Omotic language, which their forager progenitors are thought to have adopted from early Afro-Asiatic-speaking settlers in the Ethiopian highlands (Hodgson et al. (2014), Table S6). (*N.B. An earlier linkage disequilibrium-based study by Pickerell et al. (2014) posited that the West Eurasian ancestry in East Africa was only around 3000 years old and linked with the Semitic-speaking founders of the Kingdom of D’mt. Thanks to ancient DNA analysis, we now know that this biogenesis scenario is incorrect. Most of the West Eurasian ancestry in the region is actually older, higher in estimated frequency (especially when linkage disequilibrium bias is corrected for), and — as already strongly suggested by archaeological excavations — associated instead with the ancient Cushitic settlers of the Pastoral Neolithic (cf. Skoglund et al. (2017), Prendergast et al. (2018), Wang et al. (2020)).)
Afar:
69% Non-African ancestry (of which 43%=Ethio-Somali, 12%=Arabian, 14%=Others)
31% African ancestry (of which 8%=Ethiopic, 20%=Nilo-Saharan, 3%=Others)Amhara:
65% Non-African ancestry (of which 34%=Ethio-Somali, 16%=Arabian, 15%=Others)
35% African ancestry (of which 16%=Ethiopic, 16%=Nilo-Saharan, 3%=Others)Ari Blacksmith:
2% Non-African ancestry (of which 1%=Ethio-Somali, 1%=Arabian, 0%=Others)
97% African ancestry (of which 94%=Ethiopic, 1%=Nilo-Saharan, 2%=Others)Ari Cultivator:
19% Non-African ancestry (of which 17%=Ethio-Somali, 1%=Arabian, 1%=Others)
81% African ancestry (of which 63%=Ethiopic, 10%=Nilo-Saharan, 8%=Others)Ethiopian Somali:
64% Non-African ancestry (of which 53%=Ethio-Somali, 3%=Arabian, 8%=Others)
36% African ancestry (of which 8%=Ethiopic, 22%=Nilo-Saharan, 6%=Others)Morocco S:
69% Non-African ancestry (of which 44%=Maghrebi, 8%=European, 17%=Others)
31% African ancestry (of which 22%=Niger-Congo, 4%=Nilo-Saharan, 5%=Others)Oromo:
55% Non-African ancestry (of which 32%=Ethio-Somali, 12%=Arabian, 11%=Others)
45% African ancestry (of which 21%=Ethiopic, 19%=Nilo-Saharan, 5%=Others)Somali:
66% Non-African ancestry (of which 57%=Ethio-Somali, 1%=Arabian, 8%=Others)
34% African ancestry (of which 6%=Ethiopic, 23%=Nilo-Saharan, 5%=Others)Tygray:
68% Non-African ancestry (of which 35%=Ethio-Somali, 16%=Arabian, 17%=Others)
32% African ancestry (of which 12%=Ethiopic, 17%=Nilo-Saharan, 3%=Others)Wolayta:
44% Non-African ancestry (of which 27%=Ethio-Somali, 7%=Arabian, 10%=Others)
56% African ancestry (of which 35%=Ethiopic, 15%=Nilo-Saharan, 6%=Others)Although Molinaro et al. and Hodgson et al. differ on when that non-African ancestry arrived in Africa, these estimates nevertheless accord well with Molinaro et al.’s theory because the ancient Libyans (the immediate ancestors of the Maghrebans) were among the Sea Peoples.



-

- An Agaw man, representing the original Cushitic type of the Abyssinians (Habesha). Before they adopted Semitic languages, Abyssinians were Agaw speakers.
-

- An Amhara Abyssinian man, with visible South Omotic influence in terms of nasal form and hair texture. Around 17% of Amhara individuals and 10% of Ethiosemitic speakers generally carry the paternal haplogroup A, which was acquired through intermixture with South Omotic peoples.
-

- An Abyssinian woman with customary braids. The South Omotic influence can again be seen in the coarse hair texture and transverse nostrils.
-

- Although essentially of Cushitic descent, the royal family of Ethiopian Emperor Haile Selassie I, like many Abyssinians, shows both Semitic and South Omotic secondary influences. The Semitic strain is apparent in the large eyes, yellow-brown complexion and hook-nose, while the South Omotic element is clear from the shorter height and coarse hair texture.
-

- Painting of Dejazmach Hailu, Governor of Asmara (c. 1855-68). The ruler and his Abyssinian subjects are depicted with large eyes, yellow-brown complexion and coarse hair texture, as common in Abyssinian art. This reflects the Semitic and South Omotic influences on the Abyssinians.
-

- An ancient Egyptian mural depicting a Syrian Semitic man and his Egyptian wife and child (c. 1350 BCE). The Semitic-speaking natives of the Middle East were originally very close in physiognomy to the predynastic Egyptians, but gradually diverged due to increasing introgression from the Caucasus and Iranian plateau.
-

- Another ancient Semitic figure. The man has a convex nasal profile, which is very common among modern Semitic speakers. This is due to contact with earlier peoples from the Caucasus vicinity, among whom marked nasal projection, large eyes and hirsutism were distinctive attributes (e.g. the ancient Hittites; see detail on them and the Semitic-speaking Sabaeans and Himyarites on Punt: an ancient civilization rediscovered).
-

- Another ancient Semitic figure. This individual has the same overall attributes as the former, but has less pronounced nasal projection and is more deeply pigmented. He therefore likely represents the Mahra and other similar groups, who, because of their geographical isolation, have been less exposed to foreign influences. As such, they have retained more of the Semites’ original Natufian ancestry.
-

- An ancient brick wall depiction of Assyrian guards. Notice the close resemblance to the Syrian figure above. This similitude suggests that, before intermixing with lighter-skinned newcomers from the Caucasus/Iranian plateau, the Semitic inhabitants of Mesopotamia bore greater Natufian ancestry and thus were rather Mahra-like in phenotype.
-

- Statuette of an ancient Semitic male figure, found in Egypt. Note again the Mahra-like physiognomy, though less deeply pigmented due to contact with peoples from the Caucasus/Iranian plateau.
-

- Painting of the Exodus at Dura Europos, Syria, one of the earliest known synagogues (c. 245 CE). Moses (center-left) and the Israelites are depicted with the characteristic yellow-brown complexion and curly hair of the Jews and other Semitic peoples. Ancient DNA analysis of Bronze Age specimens from Sidon, Lebanon indicates that these Levantine individuals had a medium brown skin tone, as they carried the derived SLC24A5 allele (lighter pigmentation) and the ancestral SLC45A2 allele (darker pigmentation) (Haber et al. (2017)).
-

- Oldest depiction of the Iranian prophet Zoroaster (Zarathustra), founder of Zoroastrianism. Note the lighter complexion characteristic of early peoples with substantial Caucasus Hunter-Gatherer ancestry. Archaeogenetic analysis indicates that most individuals exhumed at ancient sites in Armenia, Caucasus region, had an intermediate skin pigmentation (Lazaridis et al. (2022), Supplementary Materials).
-

- Parsi singer Freddie Mercury (born Farrokh Bulsara). As an ethnic Zoroastrian, Mercury is likely close in physiognomy to the Iranian/Caucasus outsiders who originally introduced the Y-DNA haplogroup J and the Caucasus Hunter-Gatherer component to the ancestors of the Semites in the Middle East.
-

- Ancient Egyptian mural depicting the Hyksos, a Semitic-speaking people who invaded Egypt during the Second Intermediate Period (c. 1650-1550 BCE). Note the figures’ yellow-brown complexion, which is around a shade lighter than that shown on older portrayals of Semites. This is due to admixture with light-skinned newcomers from the Caucasus/Iranian plateau, which had the effect of diluting the Semites’ original Natufian ancestry. Shortly afterwards, the Hyksos would enter the Nile Valley, helping to further spread Caucasus/Iranian elements into Northeast Africa.
-

- Reproduction of ancient Egyptian wall paintings depicting Hittite (left) and Canaanite (right) figures. The Hittites were among the foreigners from the Caucasus/Iranian plateau vicinity, who genetically influenced the ancestral Semites. These newcomers introduced “Armenoid” physical traits to the Natufian-descended Canaanites, Israelites and other native Semitic peoples of the Levant, Arabia and Mesopotamia.
-

- Reconstruction of an ancient Phoenician man, with the distinctive facial features and yellow-brown complexion of the modern Levantines. The ancillary Semitic element that is present in the Abyssinians likely had a similar phenotype.
-

- Actor Adam Sandler (Askhenazi Jew). Despite some European admixture, Ashkenazi and Sephardic Jewish individuals are essentially of Levantine descent (~60%). As such, they have largely retained the physiognomy of their Semitic ancestors (cf. Atzmon et al. (2010)).
-

- Comedian Pauly Shore (Sephardic Jew). Like Ashkenazi Jews, Sephardic Jewish individuals have also largely retained the physiognomy of their Semitic ancestors.
-

- Journalist Jeffrey Toobin (Ashkenazi Jew). Ashkenazi and Sephardic Jews left their Levantine homeland and returned centuries later, with some admixture from their European host community (similar to Americo-Liberians). As such, they have largely retained the physiognomy of their Semitic ancestors.
-

- Hasidic Jewish man. Among Jewish individuals, European admixture tends to be highest among adherents of Hasidic sects, who represent a tiny portion of Jews worldwide (~5%).
-

- Actor Tomer Sisley (Mizrahi Jew). Most Mizrahi Jewish individuals have also largely retained the physiognomy of their Semitic ancestors, like their Ashkenazi and Sephardic brethren.
-

- Entrepreneur Patrick Bet David (Assyrian-Armenian). Like their Jewish and Arab kinsmen, Assyrian individuals have largely retained the physiognomy of their Semitic ancestors.
-

- Paul Newman (Ashkenazi Jew father & European mother). In German, the pejorative term “Mischling” refers to mixed individuals of Jewish and European parentage. Although such persons can be genetically differentiated from fully European individuals, they physically bear few Middle Eastern influences. In order to make their movies more commercially appealing to Western audiences, Hollywood filmmakers have, consequently, preferentially granted Jewish roles to European actors or to half-Jewish or European-passing Jewish actors. According to the director Otto Preminger (himself a Jew), this is the main reason he cast Newman in the movie Exodus (1960). In Preminger’s words, he wanted someone of Jewish heritage who did not “look Jewish,” and the naturally blond-haired, blue-eyed, half-Jew Newman fit the bill (cf. Hollywood: Where Jews Don’t Get To Play Jews).
Other examples:
-

- Actor John Turturro (Southern Italian). Historically, populations in southern Italy have had extensive contact with territories in the Middle East, particularly during the Roman period. Due to this interaction, many Southern Italians (peoples of European origin with significant Middle Eastern admixture) now overlap in physiognomy with Ashkenazi and Sephardic Jews (peoples of Middle Eastern origin with significant European admixture).
-

- Adult actor Rocco Siffredi (Southern Italian). Siffredi was born in Ortona, an area in southeastern Italy that was less impacted by Middle Eastern contact than places such as Sicily. Consequently, he closely resembles North Italian individuals.
-

- Politician Anthony Weiner (Ashkenazi Jew).
-

- Comedian Gary Shandling (Ashkenazi Jew).
-

- Saxophonist Ryan Matzinger (Ashkenazi Jew-African American).
-

- Scholar Sarah Stein (Ashkenazi Jew).
-

- Singer Liel Kolet (Mizrahi Jew).
-

- Actor Ahmed Al-Munirawi (Palestinian).
-

- Yemeni Jewish men. Because of their elevated Natufian ancestry, Yemeni Jews are the closest living representation of early Ashkenazi and Sephardic Jews, before the admixture event(s) that occurred between the latter and southern Europeans.
-

- Saudi Arab men. Note the close resemblance to the Yemeni Jewish men. This is ultimately due to shared Natufian ancestral roots.
-

- Ashkenazi Jew activist Abbie Hoffman. Observe the likeness to both the Yemeni Jewish men and Saudi Arab men. This similitude ultimately stems from shared Middle Eastern Semitic ancestral roots.

The European explorer Bertram Thomas pictured with some “pure” Arabs. The deeply pigmented, nearly black color of these individuals approximates that of the earliest Semites. This is because, prior to contact with European-related peoples bearing Anatolian Neolithic ancestry and peoples from the Iranian plateau/Caucasus bearing Caucasus Hunter-Gatherer ancestry, the ancestral Semites (who descended from the Natufians) did not bear any alleles associated with lighter coloration.
(For further details, see Are Ashkenazi Jews and Sephardic Jews really of the same ancestral origin as Mizrahi Jews, Palestinians, Lebanese, peninsular Arabs, Assyrians, Mahra and other Semites? and Why do so many Ashkenazi Jews and Sephardic Jews have afros?.)

Y-DNA affinities of populations in the Middle East, Africa and Europe. Ashkenazi Jews cluster with Syrians, Palestinians, Lebanese and other Semitic-speaking groups due to their shared Levantine ancestral origins (Hammer et al. (2000)). (*N.B. Along with their mtDNA haplogroup frequencies, the Y-DNA haplogroup frequencies of Ashkenazi and Sephardic Jews can be used to infer how much Middle Eastern ancestry and European ancestry these individuals carry. Since around 80% of Ashkenazi and Sephardic Jews bear Middle Eastern-affiliated Y-DNA lineages compared to about 20% who bear European-affiliated Y-DNA lineages, and around 20% of Ashkenazi and Sephardic Jews bear Middle Eastern-affiliated mtDNA lineages compared to about 80% who bear European-affiliated mtDNA lineages, we may surmise that, on average, Ashkenazi and Sephardic Jewish individuals carry roughly 50% Middle Eastern-associated ancestry and 50% European-associated ancestry. This corroborates genome analyses on Ashkenazi and Sephardic Jews, which have similarly found that these individuals are of Middle Eastern origin but harbor substantial European admixture (e.g. Atzmon et al. (2010)). For further details on Stefflova et al. (2009)‘s mAUTO formula, which we have used here to calculate the autosomal DNA of Ashkenazi and Sephardic Jews, see Can the Y-DNA and mtDNA of a population be used to infer its autosomal DNA?.)
-
- Y-DNA affinities of Jewish males, according to the Family Tree DNA database (Unkefer (2020)). Like Palestinians, peninsular Arabs, and other modern Semitic peoples, haplogroup J is the most frequently occurring paternal clade today among Jewish males (41%). The lineage first introgressed into the ancient Semitic gene pool around the Bronze Age, when it was introduced by foreigners from the Iranian plateau/Caucasus. This is also supported by Behar et al. (2017), who note that “haplogroup J grants the largest overall contribution to the Ashkenazi paternal gene pool, accounting for 38% of the total variation.” The next most prevalent Y-DNA haplogroup carried by Jewish males is E1b1b (20%), the main paternal lineage of the Semites’ immediate Pre-Pottery Neolithic and Mesolithic Natufian forebears. A significant minority of Jewish individuals also bear the R1 clade (~18%), a lineage that their Semitic ancestors appear to have picked up through intermixture with Indo-European speakers. All told, Jews predominantly carry Middle Eastern-associated Y-DNA lineages (~73% of the J, E, G and T clades).
-
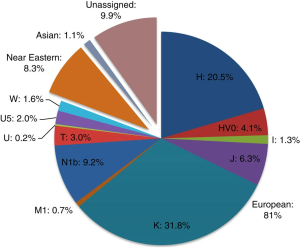
- mtDNA haplogroups carried by Ashkenazi Jews. Most Ashkenazi Jewish individuals bear maternal clades of European affiliation (~81%). This is consistent with the observation that their Middle Eastern Semitic male ancestors interbred with native women in southern Europe (Costa et al. (2013)).

In terms of autosomal DNA, Ashkenazi Jews and Sephardic Jews cluster in between other Middle Easterners and Europeans. This is a reflection of the fact that their Semitic male ancestors interbred with native women in southern Europe.

Principal Component Analysis of Askhenazi Jews and contemporary and medieval samples from the Middle East and Europe. Individuals buried at the medieval Jewish cemetery at Erfurt, Germany, form two distinct genetic clusters: one group pulls toward modern Lebanese and the other group pulls toward modern north Italians and Greeks. Ashkenazi Jews, like some Erfurt individuals, are positioned in between these two clusters. This again reflects the fact that the Semitic ancestors (mostly males) of Ashkenazi Jews and Sephardic Jews interbred with natives in southern Europe (mostly females) when they left their Levant homeland — an admixture process that was apparently still in its early stages as recently as the Middle Ages (Waldman et al. (2022)). This is also supported by linguistic analysis, which indicates that the Yiddish tongue that many Ashkenazi Jews now speak is a creole of their Levantine ancestors’ original Semitic Hebrew language and the German language of their European host community (much like how Jamaican patois is a creole of the native Niger-Congo languages of the West African ancestors of Jamaicans and the English language of their European host community). The fact that the Indo-European contact language here is a Germanic idiom spoken in west-central Europe rather than Italian or Greek conveys that the Levantine forebears of Ashkenazi Jews would still have spoken their native Hebrew language as they spread from southern Europe to other parts of the continent. (For more information, see Yes, Ashkenazi Jews (Including Gal Gadot) Are People of Color.)

STRUCTURE and phylogenetic analysis of Jewish populations. Ashkenazi Jews cluster with Mizrahi Jews from Syria and other Jewish groups. These Jewish populations carry both Levantine/Arabian (pink component) and European admixture elements (blue component). Ashkenazi individuals trace just over half of their ancestry to the Levantine/Arabian component (~60%), intermediate between Palestinians and southern European populations. This is consistent with the observation that the Semitic male ancestors of Ashkenazi and Sephardic Jews interbred with native women in southern Europe (Atzmon et al. (2010)). (*N.B. For more details, see Are Ashkenazi Jews and Sephardic Jews really of the same ancestral origin as Mizrahi Jews, Palestinians, Lebanese, peninsular Arabs, Assyrians, Mahra and other Semites?)
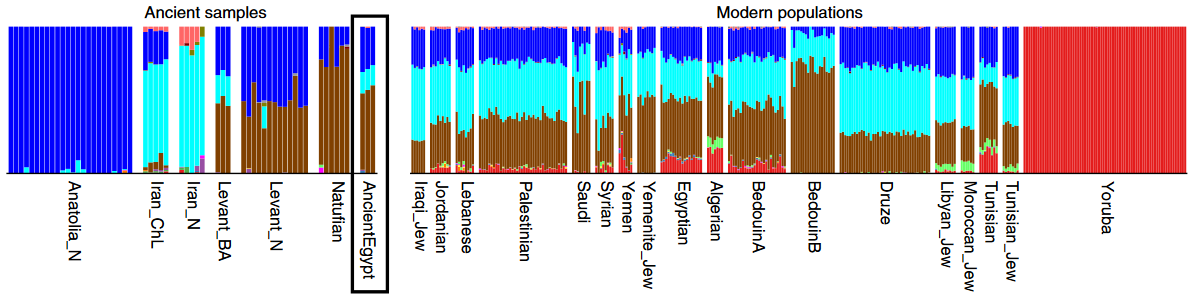
Structural genome analysis indicates that individuals belonging to the Pre-Pottery Neolithic culture of the Levant (Levant Neolithic) interbred significantly with incoming Southern European-related groups, who bore the Anatolian Neolithic ancestral component (Schuenemann et al. (2017)). Consequently, Levantine samples from this Neolithic period acquired substantial “Mediterranean” ancestry that is not pronounced among their Mesolithic Natufian ancestors. Bronze Age migrations of peoples from the Caucasus/Iranian plateau would subsequently introduce the paternal haplogroup J and the Caucasus Hunter-Gatherer/Iran Neolithic component to the ancestors of the Semitic-speaking natives of the Levant, Arabian peninsula and Mesopotamia. This would have the effect of diluting the Semites’ original Natufian ancestry as well as their Anatolian Neolithic admixture, thereby bringing them genetically closer than before to populations in the Caucasus and Iranian plateau. Modern exceptions are groups like the Mehri of southern Arabia, Yemenis of the Marib governorate, and Bedouin Arabs of the Negev Desert, who, due to their geographical isolation, have managed to retain more of the Semites’ original Natufian ancestry.

Principal Component Analysis of modern and ancient individuals. The contemporary Semitic speakers cluster with the Bronze Age inhabitants of the Levant (represented by the autochthones of Sidon; they bore the Y-DNA haplogroup E, as well as the J clade from admixture with newcomers bearing Caucasus Hunter-Gatherer/Iran Neolithic ancestry), followed by the Neolithic Levantines (Pre-Pottery Neolithic makers; they bore the Y-DNA haplogroup E and the older haplogroup CT, as well as the T and H clades from admixture with new settlers carrying Anatolian Neolithic ancestry) and the Mesolithic Levantines (Natufians; they bore the Y-DNA haplogroup E, as well as the older haplogroup CT). Ergo, despite experiencing various foreign admixtures, the modern Semites have remained genetically close to their ancient Levantine forebears (Lazaridis et al. (2016).

Genome analysis of Semitic-speaking Yemeni individuals. The Mahra/Mehri samples generally have the most Arabian ancestry (teal); four of the examined Mahra individuals almost entirely belong to that Natufian-related ancestral component. This supports the long-held belief that the Mahra represent the Semites in their “purest” form. Conversely, Yemenis from the other coastal governates of Hadramout and Shabwah have the most foreign admixture. The latter primarily consists of Sub-Saharan African elements derived from Niger-Congo-speaking West African (light green) and Nilo-Saharan-speaking East African (dark green) sources, as well as Caucasus-related admixture (peach). Additionally, the Hadramis have the most European-related admixture (lilac). This is in agreement with the theory that the ancient Himyarites of Hadramout and the adjacent southwestern provinces bore substantial Mediterranean or Sardinian-like ancestry, whereas the ancient Sabaeans in the northern territories had more pronounced Caucasus influences (see Punt: an ancient civilization rediscovered). Correspondingly, modern Yemenis from the highlands and northwest have the most Caucasus-related admixture, except for individuals in the Marib and Al Jawb governates, who have retained more of the Semites’ original Natufian ancestry (Vyas (2017).

-

- Skin reflectance values among global populations, objectively measuring average pigmentation levels. Europeans have the lightest complexions and consequently the highest skin reflectance values (over 60 at the 685 nm reading). Most African individuals have dark complexions and therefore low skin reflectance values (primarily under 40). Middle Easterners on average have intermediate complexions and thus comparable skin reflectance values (between 40 and 60). In the Middle East, Iranian peoples and Turks have the highest skin reflectance values, consistent with their elevated Anatolian Neolithic, Caucasus Hunter-Gatherer and European Steppe ancestries. Natives of Israel, a country founded and primarily inhabited by Ashkenazi Jews and Sephardic Jews, have a lower observed skin reflectance value identical to that of the autochthones of neighboring Lebanon (58.20). Since Ashkenazi Jews and Sephardic Jews carry almost 50% European admixture, this tells us that, prior to that admixture event(s), their Middle Eastern ancestors must have been similar to Mizrahi Jews and other relatively “purer” Semitic peoples (Jablonski and Chaplin (2000)).

Reconstruction of a 7500 year old Early European Farmer woman excavated in Gibraltar. Ancient DNA analysis of the specimen (named “Calpeia”) has found that she was defined by the Anatolian Neolithic ancestral component. Craniometric analysis further indicates that she and other bearers of the Anatolian Neolithic component would have had a similar “Mediterranean Caucasoid” or Sardinian-like physiognomy. However, their skin tone would likely have approached a light brown since they only possessed the derived SLC24A5 allele, the primary contributor to lighter skin pigmentation. Besides this mutation, populations in Europe now have several additional, minor light skin-associated variants, which cumulatively are responsible for the white complexion of modern Europeans (Gibraltar National Museum).
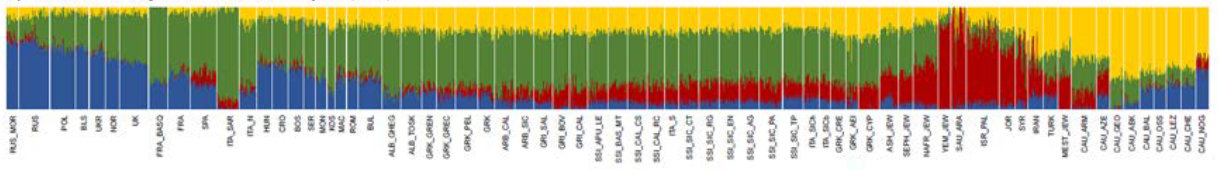
Analysis of ancient DNA samples relative to modern European, Caucasus and Near Eastern populations indicates that the Anatolian Neolithic component (green) is today most frequent among Sardinians, Basques and other southern & western European groups. The Western Hunter-Gatherer/WHG component (blue) is most common among Russians and other eastern European groups. The Caucasus Hunter-Gatherer/CHG component (yellow) peaks among Georgians and other populations inhabiting the Caucasus. The Natufian component (red) climaxes among Saudis, Yemeni Jews, Palestinians and other Semitic peoples. Ashkenazi and Sephardic Jews derive much of their ancestry from the Near East and Caucasus, with a significant admixture from southern Europe (Sarno et al. (2017), Supplementary Fig. S3). Haplogroup analysis indicates that this is because these Jewish groups’ Semitic male ancestors intermarried with native women in southern Europe when they first arrived from their Levant homeland (Nebel et al. (2000), Costa et al. (2013)).
- It helps explain the provenance of the paternal haplogroup T, which occurs at frequencies as high as 80% among Somalis. This clade is also found among more southerly Cushitic speakers, Siwa Berbers, and Tuareg Berbers, the latter of whom show especially close phenotypic ties with Somalis (see discussion below). In the archaeogenetic record, this lineage has been observed in a Pre-Pottery Neolithic B specimen excavated in the Levant (cf. Lazaridis et al. (2016), Table S6.1). Since the PPN makers are distinguished from their Mesolithic Natufian ancestors by the elevated Anatolian Neolithic admixture they bear, this suggests that the newcomers who brought the T clade to the Levant also introduced Anatolian Neolithic ancestry. Additionally, the T lineage has been detected among persons buried at Kelif el-Boroud, a late Neolithic site in Morocco. One of these ancient individuals was matched to the T-L208 (T1a1a) clade, which is ancestral to the T-Y45591 subhaplogroup that is most common among Somalis (cf. Fregel et al. (2018); YFull). According to Fregel et al. (2018), “although scarce and broadly distributed today, this haplogroup has also been observed in European Neolithic individuals” (e.g. in the Linear Pottery culture or Linearbandkeramik; see Mathieson et al. (2015)). The scientists further indicate that the Kelif el-Boroud people are largely characterized by the Anatolian Neolithic component. A southward diffusion of haplogroup T from North Africa into the Horn region is likewise supported by Sirak et al. (2021), who observed examples of both the LT parent clade and some T subclades (viz. T-L208 and T-Y31477) among Christian-era Nubian samples interred at the Kulubnarti site in Sudan (cf. Supplementary Figure 6). We have good reason to believe that LT has been in the Nile Valley for a long time since, besides its T derivative, the haplogroup L descendant lineage has also been found among ancient local specimens; Gad et al. (2020a) announced that an unidentified 18th Dynasty Egyptian royal carried the L clade. Moreover, Harney et al. (2018) report that their Chalcolithic period samples from Peqi’in Cave in Israel, which bore significant Anatolian Neolithic-related ancestry, belonged to the paternal haplogroup T. Wang et al. (2020) (discussed below) indicate that this Chalcolithic Levant sample is the best-fitting proxy for the distal source of the West Eurasian ancestry which defines the ancient Cushites of the Pastoral Neolithic.
-

- F3 statistical analysis attempting to determine the source of the Eurasian ancestry shared by the Afro-Asiatic-speaking populations in the Horn of Africa and the Toubou. Among modern groups (left), the Afro-Asiatic speakers and the Toubou showed greatest genetic affinity with Sardinians; among ancient groups (right), they showed greatest genetic affinity with the early Neolithic Linearbandkeramik farmers of Europe. Both of these populations are characterized by the Anatolian Neolithic ancestral component (Haber et al. (2016)).
-

- Y-DNA and mtDNA haplogroups of various individuals from Africa, Europe and Asia. The Toubou sample is assigned to the T1a1 paternal clade and the M1 maternal lineage, both of which are common uniparental markers among the Afro-Asiatic-speaking populations in Northeast Africa (Lorente-Galdos et al. (2019)).

Genome analysis of Daza Toubou individuals using the Vahaduo Admixture JS program indicates that they are most similar to the Kababish “Arabs” of Sudan. They share the same assortment of predominant Eurasian ancestries, which consist of majority West Eurasian elements (ancient Egyptian, European Steppe, and Levantine Natufian components) and a minority East Eurasian element (East Asian component). However, the Daza have a greater Sub-Saharan admixture than the Kababish (45.4% vs. 34.9%), as well as a more elevated Iberomaurusian admixture (14.5% vs. 0.8%).

Genetic distance analysis of modern Daza Toubou, Baggara “Arabs” and Kababish “Arabs” using the Vahaduo Admixture JS program. Almost all of these individuals show a clear preference for the medieval Kulubnarti Nubian specimens from Sudan. This confirms that the Toubou, Baggara “Arabs” and Kababish Sudanese “Arabs” share a common origin, tracing most of their proximal or recent ancestry to earlier Cushitic-related peoples.
Genome analysis of Shuwa “Arab” individuals and other modern populations from Africa. The examined Shuwa “Arabs” (labeled here “Cameroon Arabe”) derive most of their Eurasian ancestry from a Cushitic-related source (pink component), followed by a Tunisian Maghrebi-related source (light blue component). This same composite autosomal DNA signature appears to also be present at lower frequencies among the Sahelian Kanuri and Kotoko populations (who are thought to have expanded westward from the Sudan), as well as among Chadic-speaking populations of northern Cameroon (who mostly bear the Eurasian R1b-V88 paternal haplogroup). Altogether, this suggests that the Shuwa “Arabs” may originally have been Chadic speakers, who inhabited the eastern Sahara and Nile Valley beside Nilo-Saharan and Kordofanian speakers. There, through population contact, they seem to have picked up the Nilo-Saharan purple component. The Shuwa appear to have later spread to the Cameroon/Nigeria vicinity, where they settled amongst and further intermixed with Niger-Congo speakers. Such a population movement is also consistent with Basil Davidson (1968)‘s proposition that the iron-making tradition of the Nok culture in western Africa was established “perhaps by diffusion from the metal-traders of Meroe on the Middle Nile or the North African Berber states” (Bird et al. (2023)).

MSMC analysis inferring the prehistoric population size of the Toubou, Afro-Asiatic-speaking groups, and other global samples. According to the scientists, “Eurasian populations had a distinctive bottleneck at the time of their exodus from Africa ~60,000 ya. Compared to other Africans, admixed Africans (from a Eurasian gene flow), such as Egyptians, Ethiopians, and the Toubou, also showed a decline in population size during the same period” (Haber et al. (2016)).
In further support of this connection between Afro-Asiatic speakers, the ancient Sahara, and the Anatolian Neolithic component, Haber et al. (2016) indicate that the Toubou inhabitants of the Sahel carry haplogroup T at appreciable frequencies (31%). They also report that the oldest instances of the T paternal lineage have been observed among Early European Farmers from the Neolithic Linearbandkeramik (LBK) culture. Moreover, the researchers note that their sampled Toubou individuals, although admixed with neighboring Nilo-Saharan-speaking communities, are genomically more similar to the Afro-Asiatic-speaking populations in the Horn region than to local Nilo-Saharan speakers. Additionally, Haber et al. conducted f3 statistical analysis with the aim of determining the most plausible source of the Eurasian ancestry shared by these populations. They found that, among modern populations, the Afro-Asiatic speakers of the Horn and the Toubou showed greatest genetic affinity with Sardinians, and, among ancient populations, they showed greatest genetic affinity with the early Neolithic LBK farmers of Europe — both of which are characterized by the Anatolian Neolithic ancestral component. Vicente et al. (2019) also observed similar genome ties between their Toubou sample (Daza/Gorane or southern Toubou) and their Nubian and Afro-Asiatic-speaking samples from Northeast Africa. Furthermore, Haber et al. conducted an MSMC analysis to infer the prehistoric population size of their Toubou and Afro-Asiatic-speaking samples. They concluded that, like Eurasian populations that are thought to have undergone a bottleneck during the putative Out-of-Africa exodus (ca. 60,000 ybp), “compared to other Africans, admixed Africans (from a Eurasian gene flow), such as Egyptians, Ethiopians, and the Toubou, also showed a decline in population size during the same period.” They also found that “PCA of worldwide populations shows that Near Easterners and East Africans are intermediate to Eurasians and sub-Saharan Africans on PC1”, and that the Toubou individuals clustered toward the Ethiopian samples. In short, this suggests that both the Toubou and Nubians (specifically, descendants of the “red” Nubians), who now speak Nilo-Saharan languages, originally spoke languages belonging to the Afro-Asiatic family. The above securely places the earliest arrival of Minoan/LBK/Sardinian-related ancestry in Africa to the Neolithic period, thus giving a wider berth to Molinaro et al.’s thesis.
-
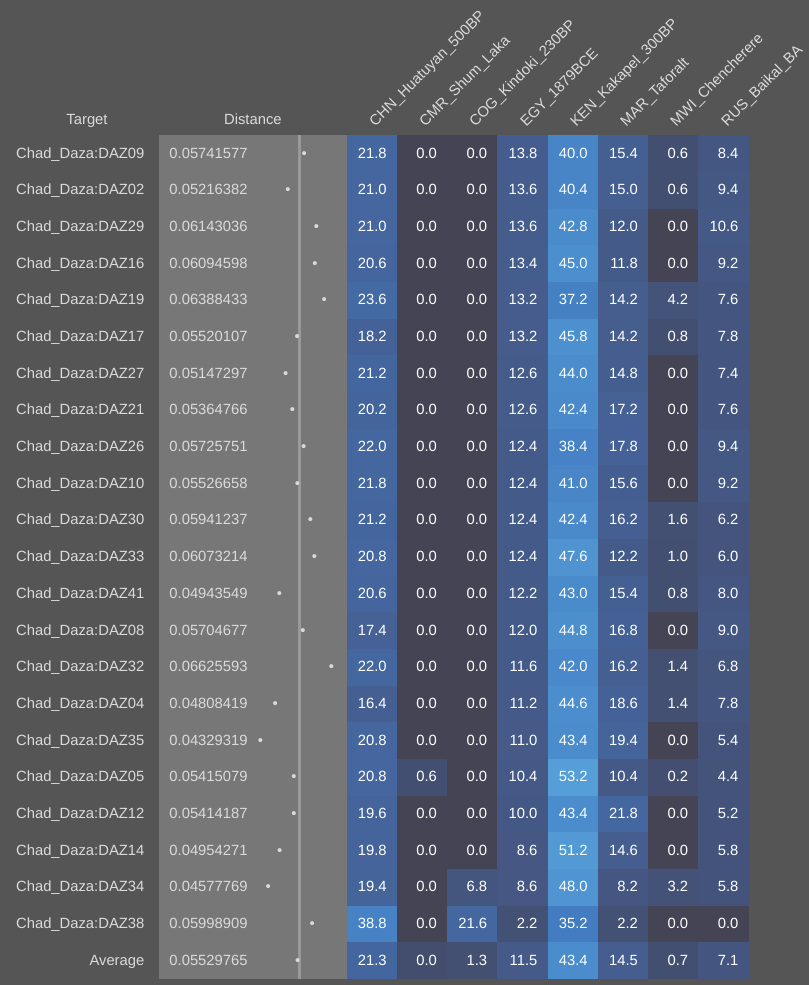



-

- A Toubou woman with the amrani/mariin phenotype. Unlike Sub-Saharan populations with significant Cushitic or Berber admixture — among whom there are no individuals with an amrani/mariin phenotype or beidan/cad phenotype — many Toubou individuals exist with the amrani/mariin phenotype. We now know from haplogroup and genome analyses that this is because they share closer genetic ties with Afro-Asiatic-speaking populations (especially the “purer” northern Toubou or Teda), and were themselves in all probability originally Afro-Asiatic speakers.
-

- A Toubou woman largely retaining the amrani/mariin phenotype, which she inherited from her Afro-Asiatic-speaking ancestors.
-

- A Toubou woman and child with the amrani/mariin phenotype. Notice the child’s soft-textured tuft of wavy hair. Such hair form is typically not found among Nilo-Saharan/Niger-Congo-speaking individuals, including those with moderate admixture from Afro-Asiatic speakers.
-

- A Toubou woman and child of relatively “pure” stock. Note again the close resemblance both physically and culturally to Afro-Asiatic speakers. This affinity has also been confirmed genetically, as the Toubou have an ancestral makeup similar to that of the neighboring Sudanese “Arabs.”
-

- Shuwa (Baggara) “Arab” women. The Baggara of Chad and Sudan are known as Shuwa “Arabs” in Cameroon and Nigeria. They are physically and genetically very similar to the Toubou, with both groups sharing affinities with Sudanese “Arabs” and other Afro-Asiatic-speaking populations. (*N.B. Earlier genome analyses of Baggara individuals suggested that they derived most of their Eurasian ancestry from source populations of either Cushitic or Berber origin. As of 2023, genome analyses indicate that they in fact carry a composite Eurasian ancestry consisting of both Cushitic and Berber components, as do Chadic speakers in northern Cameroon (albeit, at lower frequencies than the Baggara/Shuwa). As such, the Baggara/Shuwa appear to have originally spoken Afro-Asiatic languages of the Chadic branch — see Bird et al. (2023).)
-

- A Sudanese “Arab” woman. Genetic analysis suggests that the Toubou and the Baggara/Shuwa “Arabs” are a westward extension of the same, originally Afro-Asiatic-speaking ancestral stock as Sudanese “Arabs.” All of these populations trace their proximal or recent ancestry to the medieval peoples buried at Kulubnarti, Sudan, which indicates common origins.
-

Admixture analysis of Yemeni individuals. At K=3, the Yemeni samples from the Marib governorate have the most indigenous Near Eastern (Natufian) ancestry of all the examined groups, with little-to-no extraneous influences (Haber et al. (2019)).
It is in agreement with the skin pigmentation alleles that are presently found in the Horn. Researchers have observed that around 60% of the modern Afro-Asiatic speakers in the region carry the derived variant of the SLC24A5 gene, which is associated with lighter skin pigmentation (cf. ALFRED; et al. (2015)). This mutation has been detected at fixation rates (100%) among ancient Cushitic pastoralists in East Africa (Wang et al. (2020)), as well as among Minoan specimens from Crete and Kelif el-Boroud individuals. However, Natufian specimens were found to bear the ancestral SLC24A5 allele linked with darker pigmentation. The Natufians, therefore, were not the ancient population from whom the early Cushitic settlers inherited the derived SLC24A5 allele. Likewise, the Natufians do not appear to have been directly ancestral to the predynastic Egyptians. Coon (1939) indicates that many exhumed Badarian specimens were well-preserved, and their “skin was apparently brunet white, while the hair was black or dark brown in color, thick, of fine texture, and usually wavy in form.” Could perhaps, then, the derived SLC24A5 mutation have been introduced to the Cushites and Egyptians by autochthonous hunter-gatherers? It certainly was not by Ari/South Omotic peoples because the Mota specimen did not have any light skin-associated alleles. Moreover, we know from archaeogenetic analysis that during the Iron Age, Khoisan-related foragers were present as far north as coastal Kenya, where they were in contact with some Cushitic “Azanian” settlers (see Who were the ancient Azanians?). However, here too we can dismiss the Khoisan as a source of the derived rs1426654 variant at the SLC24A5 gene because a) as Lin et al. (2018) note, this “most common derived haplotype is identical among European, eastern African, and KhoeSan individuals”, b) the oldest Khoisan-related specimens analyzed (c. 4000 years old) did not carry that polymorphism (Schlebusch et al. (2017) indicate: “the SLC24A5 G allele is near fixation in African populations and all individuals with enough data exhibit the alternative G variant for the SLC24A5 gene SNP rs1426654, which codes for darker skin color”; cf. Supplementary Materials), and c) Lin et al. (2018) found evidence that the mutation was actually introduced quite recently into the Khoisan gene pool, around 2000 years ago, and likely by migrating ancient Cushitic pastoralists. Thus, it is most probable that an Anatolian Neolithic-related population (e.g. Sea Peoples, Minoan, Linearbandkeramik) and/or a Neolithic Levant-related population (Pre-Pottery Neolithic) ancestral to the early Cushites is responsible for the presence of the derived SLC24A5 variant in the Horn.

-

- Bedouins at stone ruins near Ingatat, Southern Arabia. The typically quite dark skin pigmentation of the Mahra, Socotri, Janaba, Yal Wahiba and other such “purer” Semitic populations is correlated with their elevated indigenous Natufian ancestry, for the Mesolithic Natufians did not possess any mutations associated with lighter coloration. These alleles were introduced later to the Proto-Semite natives of the Levant, Arabia and Mesopotamia: first during the Neolithic period through contact with European-related groups (bearing Anatolian Neolithic ancestry), and then during the Chalcolithic epoch via interaction with peoples from the Caucasus (bearing Caucasus Hunter-Gatherer ancestry). By contrast, the early Cushites of the Pastoral Neolithic and the ancient Egyptians were found to bear the derived variant at the SLC24A5 gene, which is associated with lighter skin pigmentation. The Natufians, therefore, were not the ancestral population from whom the Cushites and Egyptians inherited this allele. (*N.B. This has been confirmed by ancient DNA analysis. Lazaridis et al. (2022) report that most Middle-to-Late Bronze Age individuals from Israel had dark skin like their Mesolithic Natufian ancestors, whereas samples from the intervening Neolithic and Chalcolithic epochs — periods marked by an influx of newcomers bearing Anatolian Neolithic ancestry and Caucasus Hunter-Gatherer ancestry — had an intermediate complexion; cf. Supplementary Materials.)
-

- A Socotri girl. Reddish hair is found among a significant minority of Socotri, Jews and other Semitic individuals, having been recently acquired from Indo-European-speaking groups (among whom the trait appears to have originated). Red hair has also been observed among ancient Egyptian and Meroitic specimens. However, the incidence of red hair in the Nile Valley likely stems instead from older contacts with European populations (e.g., the original bearers of the paternal haplogroup R1b) since the trait has not been observed among ancient Levantine individuals dating from the Bronze Age and early periods (cf. Lazaridis et al. (2022), Supplementary Materials).
-

- Shammar Arab men in the Nejd region, Saudi Arabia. The Y-DNA haplogroup J is today the most common paternal clade borne by the Shammar Arabs and other Semitic-speaking individuals. This lineage appears to have originated in the Caucasus/Iranian plateau, from where it was later introduced to the Semites’ haplogroup E1b1b-carrying ancestors in the Levant. Consequently, the Socotri, Mahra and other south Arabian groups that have retained the most Natufian ancestry are also the Semitic peoples with the highest frequencies of the oldest haplogroup J derivatives.
-

- A Saudi Arab bedouin man retaining the deeply pigmented physiognomy of the Levantine Natufians, the ultimate ancestors of all Semites. However, the man’s sharp “Mediterranean” features are suggestive of intermixture with outsiders bearing Anatolian Neolithic or Sardinian-like ancestry. As such, he resembles a darker version of what the Pre-Pottery Neolithic culture bearers of the Levant probably looked like, since the PPN makers bore composite Natufian-Anatolian Neolithic ancestries.

South Arabian men of the Janaba and Yal Wahiba tribes. Besides the fact that the Levantine Natufians did not carry any mutations associated with lighter coloration, the idea that these ancient individuals were deeply pigmented is also suggested by the dark skin complexion of the modern Semitic peoples who carry the most Natufian ancestry. Among these “pure” Semites are the Janaba and Yal Wahiba, who have many members that are quite literally black-skinned.
-

- A Banu Hilal bedouin tribesman, North Africa. Arabian-descended peoples in North Africa and the Horn of Africa, such as the Banu Hilal and Rashaida bedouins, are frequently of a lighter complexion and more “Mediterranean” appearance than the Mahra, Socotri, Janaba, Yal Wahiba and other southern Arabian groups. We now know that this is because they carry greater Pre-Pottery Neolithic Ancestry, which was partially composed of Anatolian Neolithic (i.e. Sardinian-like) ancestry.
-

- A Somali man (Isaaq clan). Our genomic analysis has found that the Cushitic/Ethiosemitic/North Omotic-speaking peoples of the Horn of Africa carry a predominant Eurasian ancestry (~70% on average). This Eurasian ancestry consists of an ancient Egyptian component (composed of Natufian and Anatolian Neolithic elements), a European Steppe component, a Levantine Natufian component, and an East Asian component. Although Natufian elements constitute a significant portion of this Eurasian ancestry, the European Steppe, Anatolian Neolithic and East Asian components (which each have alleles embedded for lighter pigmentation) make it so that the average Afro-Asiatic speaker from the Horn is of a lighter complexion than the Janaba, Yal Wahiba and other “pure” South Arabians.

Reconstruction of a Mesolithic Natufian man, based on the Jericho Skull excavated in Israel (British Museum). Ancient DNA analysis of Natufian specimens has revealed that they were ancestral to the Levantine Neolithic population, forebears of the contemporary Semitic-speaking groups of the Middle East (Lazaridis et al. (2016)).
- It is supported by Marcus et al. (2018), who observed that the R1b-V88 haplogroup was the most common paternal lineage carried by the Middle/Late Neolithic Sardinian individuals that they examined (11/31 specimens or 35.48%). This clade is nowadays most abundant among Afro-Asiatic-speaking populations in Africa, particularly vestigial Chadic speakers in the Sahel and Siwa Berbers in the Nile Valley. The haplogroup is also borne at high frequencies by Toubou and eastern Fulani individuals. Additionally, the R1b-V88 lineage occurs at moderate percentages among Egyptian Copts, who are defined by a similar West Eurasian autosomal DNA signature as the Afro-Asiatic speakers in the Horn of Africa according to Dobon et al. (2015). Basal R1b-V88 haplotypes have been found among Mesolithic Serbian hunter-gatherers (ca. 9000 BCE), with its V2197 subclade detected among ancient Sardinian and Spanish individuals. Since the R1b-V88 lineages distributed today in Africa primarily belong to this V2197 sublineage, Marcus et al. (2020) indicate that this points to diffusion from eastern Europe to southern Europe and then to Africa of peoples carrying Early European Farmer (Anatolian Neolithic) ancestry. These population movements into Africa of Sardinian-related groups may have begun as early as the Neolithic Subpluvial (ca. 7000-3000 BCE) (cf. Supplementary Information).
Craniometric analysis of ancient pastoralists excavated at Wadi Takarkori Acacus, southwestern Libya. The Takarkori sample is most similar to Early Holocene (Kiffian culture) and Mid-Holocene (Tenerian culture) specimens from the Gobero site in the Sahara. Although chronologically younger than the Takarkori individuals, the Fezzan specimens (Fewet, Wadi-el-Ajal, Tahala) have retained a more archaic morphology and cluster instead with the late Middle Pleistocene Herto skull (Homo sapiens idaltu) (Vincenzo et al. (2015)).
Correspondingly, ancient DNA analysis of early pastoralists from Takarkori, Libya, which date from this “Green Sahara” period, were found to belong to the N maternal haplogroup (Vai et al. (2019)). Vincenzo et al. (2015) conducted cranial analysis on the Takarkori skeletal remains and observed that these individuals were most similar to Early Holocene (Kiffian culture) and Mid-Holocene (Tenerian culture) specimens from the Gobero site in the Sahara. Earlier morphological analyses report that these Gobero specimens had Iberomaurusian & Capsian and Mediterranean affinities, respectively (cf. Sereno et al. (2008); Wilford (2008)). Vincenzo et al. also found that the Takarkori individuals were physically distinct from the Fewet, Wadi-el-Ajal and Tahala specimens from the Fezzan. Although much more recent than the Takarkori sample, the Fezzan cohort had a more “archaic” morphology and clustered instead with the late Middle Pleistocene Herto skull (Homo sapiens idaltu):
The Neighbour Joining tree shows that the small sample from Takarkori takes relationships with the populations from Gobero in Niger, either coeval (Gobero B) or more ancient (Gobero A), thus with humans from sub-Saharan regions which are characterized by a wide morphological variation. Populations from the Fezzan such as Fewet, Wadi-el-Ajal and Tahala, much younger chronologically than the Takarkori sample, are separated from it (according to the length of the branches that are proportional to the Euclidean distances between the metrical variables) and are, by contrast, more in relationship with one of the earliest representatives of our species such as the cranium from Herto (Ethiopia, late Middle Pleistocene). Paradoxically, therefore, although more recent than the two women from Takarkori, samples from this time period appear more “archaic” and closer to the root of the tree.
Moreover, Haak et al. (2005) discovered that the N1a sublineage (which is common today among the Afro-Asiatic-speaking populations in Northeast Africa) was prevalent among the Early European Farmers of Neolithic Europe. This is consistent with Marcus et al.’s proposal, and also coheres well with Molinaro et al.’s suggested later in-migration of Sea Peoples.

- It reconciles the craniometric record. Brace (1993) observed that his Somali and North African samples had close biodistance values to various Neolithic populations of Europe. By contrast, Brace et al. (2006) found that their Somali and North African samples were craniometrically distant from their ancient Natufian specimens. These results support Molinaro et al.’s thesis since we know from ancient DNA analysis that the Early European Farmers of Neolithic Europe were characterized by the Anatolian Neolithic component (e.g. Lazaridis et al. (2014)). (*N.B. According to Carleton Coon (1939), the “Natufians[…] differed in physical type from period to period”, and “there was, apparently, a change of race during the Natufian.” An early period Natufian female specimen excavated at the Erg el Ahmar site had a robust skull, which metrically fit into the range of Upper Paleolithic European crania. This skeleton also “fit equally well into the North African series of Afalou bou Rummel”, a site associated with the Epipaleolithic Iberomaurusian culture. However, later Natufian skulls, such as those excavated at Shuqbah and Kebara, had a rather different, gracile Mediterranean morphology. This shift from an original Iberomaurusian-like type to a more Mediterranean type is also reflected in the archaeogenetic record. Loosdrecht et al. (2018) observed that their Iberomaurusian samples were closely related to the Mesolithic Natufians. However, Schuenemann et al. (2017) report that the Natufians had minor contact with newcomers bearing the Anatolian Neolithic component. In the ensuing Neolithic period, this embryonic Mediterranean ancestral element would grow in importance to form roughly half of the ancestry of the Natufians’ successors, the Pre-Pottery Neolithic culture bearers.)
-

- Craniometric biodistance values of Somalis, North Africans, Neolithic Europeans and other global samples. The Somalia sample is closest to the Predynastic Egypt (2.1), India (2.8), Christian Nubia (2.9), Bronze Age Nubia (3.0), Portugal Neolithic (3.1), Russia Neolithic (3.8), and Late Dynastic Egypt (3.9) samples (Brace (1993)).
-

- Craniometric analysis of ancient and modern global populations. The Prehistoric/Recent Northeast Africa lumped sample (comprising Somali, Naqada Bronze Age Egyptian, Nubian, Nubian Bronze Age, and Israeli Fellaheen Arabic farmer individuals) has closest biodistance values with the Late Prehistoric Eurasia (4.54), Algerian Neolithic (5.14), Modern Europe (5.16), Modern Mediterranean (5.22), and Prehistoric Mediterranean (5.78) samples. However, it is remote from the Natufian sample (15.31) (Brace et al. (2006)).

Craniometric analysis of various global populations. The Ethiopia sample clusters in between the Paleo Europe and modern Egypt samples (Froment (1998)).
That said, it is almost certain that there were also older population movements from and/or to the Levant. Iberomaurusian samples and early Neolithic specimens from the Ifri n’Amr or Moussa site in Morocco were found to belong to a Near East-affiliated ancestral component, which they shared with the Mesolithic Natufians and the Pre-Pottery Neolithic culture bearers of the Levant. Furthermore, these ancient specimens had no apparent Sub-Saharan affinities: Loosdrecht et. al (2018) initially posited that Taforalt had some Sub-Saharan admixture related to modern populations in West Africa. However, Lazaridis et al. (2018) subsequently found that “West Africans (represented by Yoruba) had 12.5±1.1% ancestry from a Taforalt-related group rather than Taforalt having ancestry from an unknown Sub-Saharan African source.” Lazaridis et al. (2016) also assert that “no affinity of Natufians to sub-Saharan Africans is evident in our genome-wide analysis, as present-day sub-Saharan Africans do not share more alleles with Natufians than with other ancient Eurasians.”
-

- Structural genome analysis positing ~35% Sub-Saharan African admixture in the Epipaleolithic Iberomaurusian specimens from Taforalt, Morocco. Varying degrees of similar SSA ancestry is also detected in all of the modern Afro-Asiatic-speaking samples from North Africa, the Levant, the Arabian peninsula and Mesopotamia (Serra-Vidal et al. (2019), Supplementary Information). Could this putative SSA admixture in the ancient Iberomaurusian individuals actually be a false genetic signal? According to Lazaridis et al. (2018), it is since they found that “West Africans (represented by Yoruba) had 12.5±1.1% ancestry from a Taforalt-related group rather than Taforalt having ancestry from an unknown Sub-Saharan African source.”
-

- Structural genome analysis detecting a shared West Eurasian ancestry at K=5 for the Cushites of the Pastoral Neolithic, the Epipaleolithic Iberomaurusian specimens from Taforalt, early Neolithic individuals from Ifri n’Amr or Moussa, late Neolithic samples from Kelif el-Boroud, and Chalcolithic period specimens from the Levant (Wang et al. (2020), Supplementary Material).

Genome analysis identifying a substantial North African Iberomaurusian component (orange element) in the ancestral composition of ancient Levantine individuals from the Pre-Pottery Neolithic culture of c. 8000-6000 BCE (D’Atanasio et al. (2023)).
Craniometrically, Terrazas Mata and Benavente (2013) report that no firm conclusions can be drawn on the population affinities of the Iberomaurusians. Their Taforalt sample almost exclusively grouped with other ancient individuals dating from the Middle Pleistocene, Upper Paleolithic and Early Holocene, and therefore in their judgement “these populations are still too far away from the Neolithic and Historic samples, so they could not represent direct ancestors of any present human population.” However, their modern Horn of Africa and Dynastic Egypt samples clustered together. The scientists indicate that this is because “these groups seem to represent migrations of groups from the Middle East, who spoke languages of the Afro-Asiatic family”. (*N.B. Terrazas Mata and Benavente state that “only two recent populations, both from Western Africa, have been integrated into the third branch, being near to the Paleolithic sample from Taforalt, Algeria”. However, the scientists do not explain the reason behind this apparent affinity. It may be that Terrazas Mata and Benavente’s Taforalt sample contains some admixed individuals since Brace et al. (2006) observed that their Taforalt/Afalou cohort clustered with Upper Paleolithic European crania rather than with any of their Sub-Saharan samples. Another possibility is that this is a manifestation of gene flow in the other direction, from the Iberomaurusians into the Proto-Niger-Congo population. As suggested above (under Final observations and recommendations), the Iberomaurusians may have introduced the paternal haplogroup E to adjacent Sub-Saharan African groups, a clade of which they and the related Natufians are now known to be the oldest carriers. This admixture event(s) probably also transmitted some West Eurasian genes, which Lazaridis and other researchers have detected in their Yoruba and other modern West African samples. Alternatively, a population(s) with considerable Iberomaurusian ancestry (e.g. the ancestral Fulani) may have been responsible for spreading the E lineage to the Niger-Congo-speaking peoples. The Niger-Congo speakers may instead have originally borne the archaic A and B haplogroups given: 1) the young TMRCA of the E1b1a or E-M2 subclade, which most Niger-Congo-speaking individuals today carry (estimated at just 6175-6588 years before present; cf. Ansari Pour et al. (2013)), 2) the genomic ties between Niger-Congo speakers and the primary carriers of the A and B lineages (viz. Khoisan, Pygmies, Nilo-Saharan speakers, Hadza and other East African hunter-gatherers), 3) the disconnect between the autosomal DNA of the Niger-Congo-speaking populations and those of the other haplogroup CT-descended global populations, 4) the fact that, with the exception of the Mota sample (which has trace levels of Eurasian admixture — see Erratum above), all of the pre-Bantu Migration ancient specimens in Sub-Saharan Africa that have thus far been genetically analysed belong to the archaic A and B paternal haplogroups (Lipson et al. (2022), Extended Data Table 1), and 5) the earliest and exclusive presence of the CT and DE parent clades among ancient specimens in Asia.)
-

- Y-DNA haplogroups carried by various ancient individuals in Sub-Saharan Africa. Except for Mota (which has trace levels of Eurasian admixture), all of the pre-Iron Age specimens analyzed belong to the archaic A and B paternal lineages. This supports the view that the modern Niger-Congo-speaking populations originally bore the A and B clades and acquired the E haplogroup only recently; either directly from early Afro-Asiatic speakers or indirectly via the Bantu Migration of ~2000 ypb (Lipson et al. (2022), Extended Data Table 1).
-

- Archaeogenetic analysis conducted thus far indicates that the oldest instances of the archaic Y-DNA haplogroup A are found among specimens in Southern Africa, whereas the oldest examples of the paternal clade E have been found among North African and Middle Eastern specimens (Martiniano et al. (2022)).
Ultimately, the Iberomaurusians appear to have originated from outside Africa; most probably somewhere near Europe given their craniometric and mtDNA ties with Upper Paleolithic Europeans. Accordingly, the oldest instance of U6*, a signature maternal haplogroup borne by Iberomaurusian specimens, has been observed in Peştera Muierii 1 (PM1), a 35,000 year old individual excavated in Romania (Hervella et al. (2016)):
After the dispersal of modern humans (Homo sapiens) Out of Africa, hominins with a similar morphology to that of present-day humans initiated the gradual demographic expansion into Eurasia. The mitogenome (33-fold coverage) of the Peştera Muierii 1 individual (PM1) from Romania (35 ky cal BP) we present in this article corresponds fully to Homo sapiens, whilst exhibiting a mosaic of morphological features related to both modern humans and Neandertals. We have identified the PM1 mitogenome as a basal haplogroup U6*, not previously found in any ancient or present-day humans. The derived U6 haplotypes are predominantly found in present-day North-Western African populations. Concomitantly, those found in Europe have been attributed to recent gene-flow from North Africa. The presence of the basal haplogroup U6* in South East Europe (Romania) at 35 ky BP confirms a Eurasian origin of the U6 mitochondrial lineage. Consequently, we propose that the PM1 lineage is an offshoot to South East Europe that can be traced to the Early Upper Paleolithic back migration from Western Asia to North Africa, during which the U6 lineage diversified, until the emergence of the present-day U6 African lineages.
Furthermore, this part of Europe, the Balkans, remains a hub of V13, the most prevalent Europe-centered derivative of the E1b1b paternal haplogroup. The Iberomaurusians carry E1b1b’s M78 sublineage, which is the parent clade of V13. There are, therefore, also Y-DNA ties. In short, since the ancient West African-related population that the Iberomaurusians are hypothesized to have had contact with was indigenous to Africa, it follows that the Iberomaurusians must have lacked any such admixture when they first settled there. Future testing of older Iberomaurusian specimens is thus likely to yield examples of relatively unadmixed individuals. (For further details, see Did the Capsians of North Africa descend from the Iberomaurusians?, What are the genetic affinities of the Aterians?, and Have the Kiffians and Tenerians of the Gobero been genetically analysed yet?.)
-

- Reconstruction of an Epipaleolithic Iberomaurusian man. Along with the Mesolithic Natufians of the Levant, the Iberomaurusians of North Africa have been found to be the oldest bearers of the paternal haplogroup E (Science Source).
-

- Reconstruction of the Nazlet Khater specimen, a Paleolithic skeleton of c. 30000 BP excavated in Egypt. Biometric analysis indicates that Nazlet Khater is morphologically distinct from ancient and modern North African specimens (including the Iberomaurusians of Taforalt), and is instead most similar to Sub-Saharan Late Middle Stone Age specimens. According to Vermeersch (2002), this suggests that Nazlet Khater represents a “Proto-Khoisan” stock that once inhabited areas as far north as the Nile Valley. This artistic reconstruction, with its resemblance to the modern Khoisan, supports that theory. However, it is incorrect in two important respects. Firstly, the Proto-Khoisan would have had a darker complexion. We know this because the derived rs1426654 allele at the SLC24A5 gene, which is the primary light skin-conferring allele that contemporary Khoisan individuals bear, was transmitted to their ancestors by ancient Cushitic pastoralists (Lin et al. (2018)). Older Khoisan specimens (c. 4000 BP) dating from before that contact period did not carry that mutation (Schlebusch et al. (2017), Supplementary Materials). Secondly, scientists have observed that the distinctive “narrow eye opening” and “peppercorn” hair of Khoisan individuals appear to be controlled by the rs111298318 allele of the KRT74 gene. Fan et al. (2023) report that this rs111298318-C mutation is carried by most Khoisan samples they analysed (73%), whereas it is rare among other populations of Africa and Eurasia (5%). Thus, the reconstruction, although generally accurate in terms of facial structure, should actually show a darker-skinned Nazlet Khater specimen with “peppercorn” hair.
-
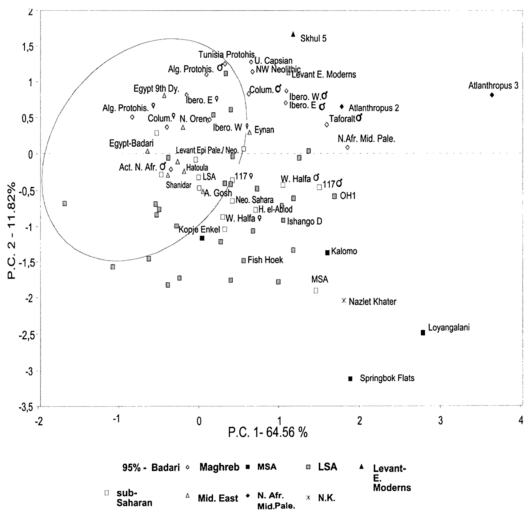
- Craniometric analysis of the Nazlet Khater specimen and other global ancient and modern individuals. The Nazlet Khater cohort clusters nearest to the Sub-Saharan Late Middle Stone Age specimens (MSA), and more distantly with ancestral Khoisan samples from Loyangalani, Springbok Flats, Kalomo and Fish Hoek. By contrast, the Nazlet Khater specimen is morphologically distinct from the Predynastic Egyptian Badarian sample, which groups instead with modern North African individuals (Actual North Africa), Algeria Protohistoric individuals and other ancient North African and Levantine specimens. Nazlet Khater is also distinct from the Taforalt and other Iberomaurusian samples. The latter individuals cluster instead with the Capsians and Neolithic Northwest African specimens, who in turn are nearest to the modern Levant sample (Vermeersch (2002)).
Moreover, the Iberomaurusians of Taforalt, the Ifri n’Amr or Moussa specimens, the Pre-Pottery Neolithic makers and the Natufians all bore the E1b1b paternal clade. This makes them, along with a 12,000-8,000 BCE Mesolithic specimen from the Belt Cave in Iran (Narasimhan et al. (2019)), the oldest known carriers of the haplogroup E. Some Natufian individuals were also found to carry the Y-DNA haplogroup CT, which is ancestral to the DE (parent clade of haplogroups D and E) and CF (parent clade of haplogroups C and F) lineages. Additionally, multiple other instances of both CT and DE have been observed among ancient specimens in Asia, whereas old examples of these lineages have yet to be observed anywhere in Africa (cf. Narasimhan et al. (2019), Table S1-S5). Since these are the earliest and only instances of these clades in the archaeogenetic record, this strongly bolsters the argument that Asia rather than Africa is the true point of origin and dispersal of the CT and DE haplogroups. These early peoples likewise all harbored maternal lineages derived from the M and N mtDNA macroclades. Altogether, this suggests that, as in the case of the paternal haplogroup T, the contemporary Afro-Asiatic speakers in the Horn of Africa and North Africa inherited their E1b1b sublineages from ancestors with a Eurasian genetic background (Fregel et al. (2018); Loosdrecht et al. (2018); Lazaridis et al. (2016)).
African admixture component
When the Mota man was excavated in Ethiopia, some scholars wondered if the population to which he belonged might represent the native hunter-gatherers whom the Afro-Asiatic-speaking agropastoralists encountered and possibly mixed with when the latter first settled in the Horn. More extensive DNA analysis has, however, shown that gene flow between the two communities, although present, was minimal (Lipson et al. (2020) places Mota-related admixture in the Cushitic-speaking Agaw at 8%; no other hunter-gatherer-related admixture has been detected in the Horn). Due to this apparent lack of a suitable ancient proxy sample, researchers have formulated alternative admixture scenarios using various modern populations. In qpAdm and other genomic programs, these contemporary reference groups have served as stand-ins for the elusive ancient African contact population(s). The Dinka, Nuer and other northern Nilotic communities have often been the go-to populations utilized as the African proxies. The problem is that all of these groups have proven to be even less reliable surrogates than Mota.
Let us examine why that is:
-
- The Dinka, Nuer, Shilluk and other northern Nilotes are not ‘purely’ African populations to begin with. Like the Nilote groups to their south in the Horn (e.g. Kunama, Nara, Gumuz and Mursi) and Great Lakes (such as the Maasai, Kalenjin, Samburu and Turkana), the northern Nilotes also have considerable non-African admixture. This was already suggested by the uniparental lineages that they carry. Hassan et al. (2008) observed that around 20% of Shilluk and 15% of Dinka individuals bear the E1b1b/E3b haplogroup, a paternal clade that is most common among Afro-Asiatic speakers. Most strikingly, Balemi (2018) reports that 51.62% of his Nuer sample from southwestern Ethiopia carry the E1b1b-M78 haplogroup, compared to 16.67% of Hassan et al. (2008)’s Nuer sample from Sudan. Furthermore, 55.55% and 33.33% of Balemi (2018)’s Berta and Gumuz samples, respectively, bear the Eurasian F-M89 clade. In Africa, the earliest instance of this lineage has been observed among Christian period specimens entombed at sites on Meroë Island and the Nile’s 4th cataract (Yousif and Eltayeb (2009)). (*N.B. After the publication of Yousif and Eltayeb (2009)’s study, it was recognized that the M282 mutation, then classified under F-M89, had been misclassified. M282 was subsequently reassigned to its proper lineage, the paternal haplogroup H; specifically, to the H2 sublineage. The oldest known instance of H2 in Africa has been detected in an ancient Egyptian specimen, the Middle Kingdom aristocrat Nakht-Ankh (cf. HaploTree). It is therefore likely that this Eurasian clade was indeed introduced into the Nilotic gene pool by Afro-Asiatic speakers.) Non (2010) likewise notes that over 40% of her Sudanese Nuer sample belongs to the mtDNA haplogroups M and N, including around 18% M1 carriers (cf. Table 3-3). The M1 subclade has been found in ancient Maghreban, Egyptian and South Cushitic specimens, and still remains a signature maternal lineage among the modern Afro-Asiatic-speaking populations in Northeast Africa. In short, uniparental markers indicate that there was significant gene flow from Afro-Asiatic-speaking groups (especially Cushitic speakers) into neighboring Nilote communities.
By contrast, in various autosomal DNA studies, the northern Nilotes have appeared to be almost completely of African ancestry. For example, in the African Genome Variation Project’s analysis, the Dinka sample showed no extraneous influence at K=2 (cf. Gurdasani et al. (2015)). This apparent lack of non-African affinities is why the Dinka, Nuer and other northern Nilotes have often been used by researchers as proxies for the African component. In 2017, Skoglund et al. compared the same AGVP Dinka sample to that of an ancient South Cushitic pastoralist (Luxmanda), the first such specimen to be genetically analyzed. In their admixture analysis, the Dinka individuals now all of a sudden showed almost 30% non-African ancestry at the K=2 level. Judging by the existing uniparental marker data, it’s pretty clear why that is: there was non-African ancestry buried within the northern Nilote gene pool, and that ancestry was specifically derived from earlier Cushitic peoples such as Luxmanda.This, in a nutshell, explains why the Dinka, Nuer and other northern Nilotes often fit well as proxies for the African component in admixture testing models. However, the problem with using such admixed groups in these analyses is that doing so leads to highly distorted estimates of ancestry proportions. For example, let’s say a researcher is testing admixture models on qpAdm and finds that his modern Cushitic or Ethiosemitic-speaking population is best modeled as 50% Neolithic Levantine + 50% Dinka. What that program is really telling him is that his sample is best modeled as 50% Neolithic Levantine + X% ancient Nilotic + X% ancient Cushitic. This again stems from the fact that the Dinka are not a purely African population, but rather a Nilote-descended community with significant Cushitic admixture (whence was derived their non-African ancestry). When factoring in the ~30% of non-African ancestry that Skoglund et al. (2017) detected in their Dinka reference sample, the estimated whole genome ancestry apportionment then actually becomes 50% Neolithic Levantine + 35% African + 15% unclassified Eurasian. This necessary adjustment therefore brings the non-African total to around 65%. The same corrective adjustment would have to be made if the African reference sample were a southern Nilotic group, such as the Maasai Nilotes (e.g. when the ~30% of West Eurasian admixture in Ali et al. (2020)‘s East African proxy sample (Maasai) is taken into account, the estimated non-African ancestry for their northeastern Somali sample rises to 70%; this more accurate total is close to Hodgson et al.’s 66% average for their general ethnic Somali sample). That’s also before correcting for linkage disequilibrium bias, which, as Hodgson et al. (2014) observed and Choudhury et al. (2020) also demonstrated, would increase the Cushitic speakers’ Eurasian total even further. Put simply, there is considerable extra non-African ancestry in the genome of the Afro-Asiatic-speaking sample which is not being counted. This results in an inaccurate overall estimation of ancestry proportions. It also conflicts with scientific data gathered through other means (viz. craniometric analysis, anthropometric analysis, linguistics).
-

- Genome analysis detecting almost 30% non-African ancestry in a sample of Dinka. This Nilotic population was previously assumed to have little-to-no non-African ancestry, and was therefore often used as a proxy for inferring African ancestry (Skoglund et al. (2017)).
-

- Genome analysis again detecting moderate non-African admixture (~20%) in a sample of Dinka individuals (Scheinfeldt et al. (2019)).
-
- Genome analysis again detecting notable non-Sub-Saharan African admixture in Nilo-Saharan-speaking samples from the Nile Valley and Sahel. In this study, most of that foreign gene flow is proposed to have been derived from groups related to the Iberomaurusians, a North African people of the Epipaleolithic (D’Atanasio et al. (2023)).
-
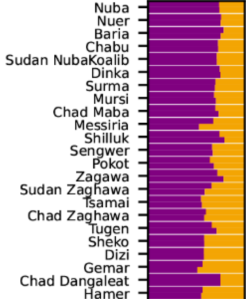
- Genome analysis once more detecting a notable non-African admixture in the Dinka, Nuer, Nuba and other Nilo-Saharan-speaking individuals from the Nile Valley and elsewhere (yellow component) (Pfennig et al. (2023)).
-
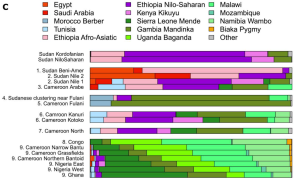
- Genome analysis again detecting non-Sub-Saharan African admixture in Nilo-Saharan and Kordofanian-speaking samples from Sudan. This pink component, which the scientists note has Arabic-related affinities, is maximized among Somalis. It is found at low-to-moderate frequencies among the examined Nilo-Saharan and Kordofanian-speaking individuals (~20%). Hence, Nilo-Saharan/Kordofanian-speakers bear Eurasian-related admixture, which was specifically acquired from Cushitic peoples (Bird et al. (2023)).
-

- Y-chromosome analysis of Dinka males has found that a significant minority bear the E1b1b haplogroup (4 out of 16 examined individuals or ~15%). This is a common paternal lineage carried by Afro-Asiatic speakers, which further underlines the fact that the Dinka and other northern Nilotes do have some non-African admixture (Hassan et al. (2008)).
-
- The Dinka, Nuer, Shilluk and other northern Nilotes are not ‘purely’ African populations to begin with. Like the Nilote groups to their south in the Horn (e.g. Kunama, Nara, Gumuz and Mursi) and Great Lakes (such as the Maasai, Kalenjin, Samburu and Turkana), the northern Nilotes also have considerable non-African admixture. This was already suggested by the uniparental lineages that they carry. Hassan et al. (2008) observed that around 20% of Shilluk and 15% of Dinka individuals bear the E1b1b/E3b haplogroup, a paternal clade that is most common among Afro-Asiatic speakers. Most strikingly, Balemi (2018) reports that 51.62% of his Nuer sample from southwestern Ethiopia carry the E1b1b-M78 haplogroup, compared to 16.67% of Hassan et al. (2008)’s Nuer sample from Sudan. Furthermore, 55.55% and 33.33% of Balemi (2018)’s Berta and Gumuz samples, respectively, bear the Eurasian F-M89 clade. In Africa, the earliest instance of this lineage has been observed among Christian period specimens entombed at sites on Meroë Island and the Nile’s 4th cataract (Yousif and Eltayeb (2009)). (*N.B. After the publication of Yousif and Eltayeb (2009)’s study, it was recognized that the M282 mutation, then classified under F-M89, had been misclassified. M282 was subsequently reassigned to its proper lineage, the paternal haplogroup H; specifically, to the H2 sublineage. The oldest known instance of H2 in Africa has been detected in an ancient Egyptian specimen, the Middle Kingdom aristocrat Nakht-Ankh (cf. HaploTree). It is therefore likely that this Eurasian clade was indeed introduced into the Nilotic gene pool by Afro-Asiatic speakers.) Non (2010) likewise notes that over 40% of her Sudanese Nuer sample belongs to the mtDNA haplogroups M and N, including around 18% M1 carriers (cf. Table 3-3). The M1 subclade has been found in ancient Maghreban, Egyptian and South Cushitic specimens, and still remains a signature maternal lineage among the modern Afro-Asiatic-speaking populations in Northeast Africa. In short, uniparental markers indicate that there was significant gene flow from Afro-Asiatic-speaking groups (especially Cushitic speakers) into neighboring Nilote communities.
- Genetic research has found that a significant portion of the core Sub-Saharan ancestry of the northern Nilotes (Dinka, Nuer, Shilluk, etc.) is related to West African populations (e.g. Skoglund et al. (2017)). However, Prendergast et al. (2008) remark that there was “no evidence of western African-related ancestry” in the ancient Cushitic individuals that they examined. Skoglund et al. (2017) and Wang et al. (2020) likewise did not detect any West African affinity in the early Cushitic specimens that they analyzed. This is hardly surprising since West African-related ancestry appears to have arrived in East Africa much later, during the Iron Age with the first Bantu settlers of the Great Lakes region. A specimen excavated at the Deloraine Farm in Kenya, the Rift Valley’s oldest agricultural site, is the earliest individual found to carry such West African-related ancestry (dated ca. 1170-970 BP; cf. Prendergast et al. (2008)). This makes modern Nilotic individuals unrealistic proxies for inferring an African admixture component among the ancient Cushitic settlers of the Pastoral Neolithic.
-

MtDNA (maternal) haplogroup affinities of various populations in Africa and Asia. The Afro-Asiatic-speaking Somali, Oromo, Afar, Tigrai and Gurage samples cluster with Yemeni individuals of Hadramout, as well as the Dawoodi Bohra and other Muslim groups of South Asia that originated from Arabia. The Dinka, Nuer and Datog Nilote samples cluster instead with other Nilotic, Bantu and hunter-gatherer populations of East Africa (Non (2010)). (*N.B. Mohamoud (2006) observed similar affinities in his analysis of HLA antigens, noting that “the result of HLA class I and class II antigen frequencies show that the Somali population appear more similar to Arab or Caucasoid than to African populations”.)
The maternal haplogroups distributed today among the northern Cushitic and Ethiosemitic-speaking populations in the Horn of Africa do not support the claim that there was significant admixture between their Afro-Asiatic-speaking male ancestors and Nilotic females. This idea first gained prominence after the publication of Watson et al. (1996) and Watson et al. (1997), two early mtDNA studies which reported that the analyzed Somali (sampled in Kenya) and Tuareg Berber (sampled in Niger) individuals primarily belonged to the L3 mtDNA haplogroup. Comas et al. (1999) similarly indicated that only 5%-27% of Ethiopians bore non-African mtDNA lineages. However, it turns out that these results were inaccurate since the L3 macroclade’s phylogeny had not yet been fully resolved. Specifically, many of the samples that were at the time presumed to represent L3 carriers actually belonged to derivatives of the N haplogroup. Martin Richards, who served as a co-author on Watson et al. (1997), explains the situation thusly:
The analysis of Watson et al. (1997) was based largely on control-region sequence data, which fails to resolve many mtDNA haplogroups. By targeting newly identified coding-region variants, Quintana-Murci et al. (1999) distinguished two major clades in non-Africans, within haplogroup L3. One of these had already been identified as the Asian super-haplogroup M (Torroni et al. 1994a); Quintana-Murci et al. (1999) showed that all other non-African L3 lineages fell into a second major clade, later named haplogroup N[…] haplogroup M was present at high frequency in Ethiopia and Somalia
Non (2010) later retested Watson et al.’s Somali sample, in addition to Oromo, Afar, Amhara and Tigray samples from Ethiopia. She found, given the now better understood mtDNA phylogeny, that around 60% of the examined individuals actually bore the Eurasian M and N haplogroups. Ottoni et al. (2009) likewise reanalysed Tuareg individuals and noted that the most common mitochondrial haplogroup among them was, in fact, the H1 lineage, which is associated with Europe. Subsequent mtDNA studies by Kivisild et al. (2004), Holden (2005) and Mikkelsen et al. (2012) observed comparable haplogroup frequencies for these Afro-Asiatic-speaking populations, including a very low occurrence of the L5 clade (<1% on average). This is of particular importance since the L5a subclade is the most common maternal lineage among the Dinka Nilotes in the Sudan area; 31% of Dinka individuals there belong to this subhaplogroup (cf. Hassan et al. (2009)). Thus, instead of showing appreciable Nilotic affinities, the mtDNA profiles of the northern Cushitic and Ethiosemitic-speaking populations in the Horn region parallel those of certain other Afro-Asiatic-speaking communities in the Arabian peninsula and North Africa (e.g. Non (2010) observed that her Somali sample clustered with a Yemeni sample from Hadramout, Kujanova ́ et al. (2009) note that 68.6% of Egyptians from El-Hayez oasis carry M and N lineages, Bekada et al. (2015) report a similar frequency of ~63% M and N haplogroups among their Algerian cohort from Algiers (see S5 Table), Coudray et al. (2009) likewise reveal that 55.3% of Figuig Berbers from northern Morocco are M and N carriers, and Kivisild et al. (2004) indicate that 56.4% of their Yemeni sample bears M and N derivatives.) These findings are why Shriner et al. (2016) propose that African introgression may have primarily occurred through the paternal line via the E1b1b haplogroup rather than through the “Arabian” maternal line. However, this argument, too, is no longer tenable because ancient DNA analysis has demonstrated that the haplogroup E was first affiliated with Epipaleolithic & Neolithic North African and Middle Eastern groups rather than Nilo-Saharan-speaking populations. The early Nilo-Saharans instead appear to have originally belonged to the Y-DNA haplogroup A (see Ancient DNA from Sudan).

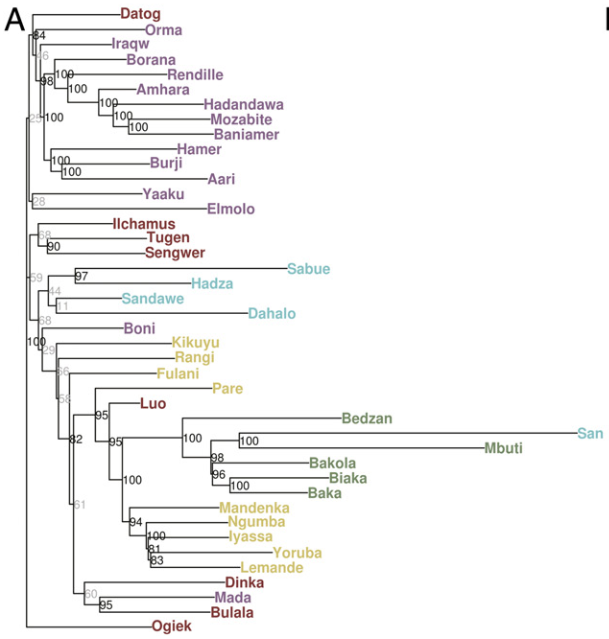
Population tree estimating Fst genetic distance between ethnic groups in Africa. The Cushitic Beja samples (Beni Amer & Hadendoa) cluster with the Mozabite Berber sample. Despite some recent Arabian and Sub-Saharan contacts, this suggests that the Beja have retained much of their Cushitic ancestry (as also observed by Dobon et al. (2015) and Gopalan et al. (2019)). These continued ties ultimately reflect the northern origin of the early Cushitic settlers of East Africa (Scheinfeldt et al. (2019)).
- According to Hassan et al. (2009), the Beja, Nubian and Sudanese “Arab” individuals that they examined almost exclusively belonged to derivatives of the mtDNA macrohaplogroup L (almost all of which were haplogroup L3 subclades). Since L is the most common mitochondria today in Africa, this observation has sometimes been used to buttress the claim that the Proto-Cushites possessed considerable Sub-Saharan admixture. However, we have good reason to doubt that this was the case at all. Firstly, Non (2010) examined other Nubian individuals and found that many of them instead bore M and N maternal lineages, and at a comparable frequency as the Afro-Asiatic-speaking samples from the Horn of Africa to boot: Tigrai=63.6%, Somali=60%, Afar=56.2%, Gurage=55%, Oromo=53.3%, Nubian=48%, Amhara=46%. This leaves open the possiblity that the Beja, like Nubians, were undersampled too, and that many of them do in fact carry M and N mtDNA lineages. Secondly, as discussed below, the oldest Cushitic settler in East Africa thus far examined actually belongs to a European Neolithic-associated mitochondrial haplogroup. Only later do Sub-Saharan-affiliated clades begin to appear in earnest, as the Cushitic pastoralists are slowly assimilated into neighboring hunter-gatherer/Nilo-Saharan/Bantu communities.Thirdly, although many modern Beja have to a remarkable extent retained the physiognomy of the earliest depicted Beja (and Cushitic) mural figure, other Beja individuals have clearly been impacted by recent contacts with Sub-Saharan populations. We can be pretty confident of this since Gibreel et al. (2018) observed an elevated frequency of sickle cell anemia among their Beja samples from Sudan (Beni Amer=22%, Hadendoa=11%). The scientists ascribe this to intermixture with the Hausa, Fulani and other local groups of West African origin because a) these populations have a high frequency of the sickle cell gene (30% among the Hausa), and b) all of the sickle cell haplotypes they observed in their Beja and other Sudanese samples belong to the West Africa-specific Benin haplotype. By contrast, Mohamed et al. (2021) did not detect any sickle cell variants among their examined Beja individuals. Foy et al. (1954) and Aluoch et al. (2006) similarly indicate that none of their modern Cushitic-speaking Somali and Oromo samples carried the sickle cell gene. Likewise, Prendergast et al. (2018) report a complete absence of sickle cell variants among the ancient Cushitic specimens they examined (cf. Table S12). Given these established contacts between certain Beja groups and West African populations, it is not unexpected that the Beja should have a greater frequency of Sub-Saharan mitochondrial lineages than other Afro-Asiatic-speaking groups in Northeast Africa.
- Ancient DNA analysis of Cushitic individuals of the Pastoral Neolithic cultural complex (discussed below) has found that they bore the derived alleles at the SLC24A5 locus (2 out of 2 examined specimens or 100%) and APBA2 locus (2 out of 2 examined specimens or 100%), which confer lighter skin pigmentation. Some individuals also carried the derived rs1800404 variant at the OCA2/HERC2 locus, an allele associated with lighter eye color (2 out of 2 examined Pastoral Neolithic specimens from the Lukenya Hill site). Additionally, at least one of the specimens carried the C-14010 mutation linked with lactase persistence. These alleles are today still common among Cushitic-speaking groups. By contrast, the early Nilotes of the Pastoral Iron Age were found to bear the ancestral alleles at the SLC24A5, APBA2 and OCA2/HERC2 loci, which are associated with darker skin pigmentation and eye color; they also lacked any derived lactase persistence alleles, including the C-14010 variant (Wang et al. (2020), Table S7). Modern Nilotic individuals in the Nile Valley likewise do not carry these derived alleles (cf. Crawford et al. (2017); Hollfelder et al. (2020)). This suggests two things: 1) ancient Nilotic populations did not significantly contribute to the gene pool of the early Cushites, and 2) gene flow from ancient Cushitic peoples toward early Nilotic groups (which has been confirmed by haplogroup and genome analyses; see above) must have started before the Neolithic era. That is, at a time period when the Cushites’ ancestors were still Iberomaurusian/Natufian-like and had not yet evolved/acquired these derived alleles.
- The linguist Alexander Militarev (1984) identified a Cushitic substratum in the Modern South Arabian (MSA) languages. He asserts that this demonstrates that Cushitic peoples originally inhabited the Arabian peninsula alongside Semitic speakers, from where most would later depart to populate Northeast Africa (cf. Blažek (2013)):
Having identified a Cushitic-like substratum in Modern South Arabian, Militarev (1984, 18-19; cf. also Belova 2003) proposes that Cushites originally lived throughout the Arabian Peninsula; thus they would be the original southern neighbors of the Semites, who then assimilated those Cushites who did not move into Ethiopia. This hypothesis is supported by Anati (1968, 180-84), who analyzed the rock art of Central Arabia. He connected the pictures of the ‘oval-headed’ people depicted with shields with the Arabian ‘Cushites’ from the Old Testament [Genesis 10.6-12; Isaiah 45.14] described also with specific shields [Jeremiah 46.9; Ezekiel 38.5]. The spread of Cushites in Africa is connected with the Rift Valley. In the coastal area of Eritrea and Djibuti, where the Rift enters into the African mainland, three archaic representatives of the North, Central (= Agaw) and Eastern branches of Cushitic appear: Beja, Bilin and Afar-Saho respectively. In this place the disintegration of Cushitic probably began.
Vyas (2017) also genetically analyzed Mehri (Mahra) individuals from Yemen’s Mahra governorate, who natively speak Mahra, a Modern South Arabian language. He found that, in contrast to other Arabian and Levantine populations, his Mehri sample experienced minimal exogenous admixture:
While admixture and gene flow generally characterize the Levantine and southern Arabian populations in my studies, some groups have been found to be minimally admixed. When I studied the ancestral components present in Levantine and southern Arabian populations, I found that many of the Yemeni from the Mahra governate were largely non-admixed and (almost) all of their ancestry derived from the Arabian IAC. All the sample donors from the Mahra governate reported Mehri (i.e., a Modern South Arabian language spoken in parts of Yemen and Oman) as their primary language; the people of Mahra are thus linguistically isolated from the neighboring Arabic-speaking groups (Rubin 2008; Rubin 2010).
Had there been a pronounced Sub-Saharan influence among the Proto-Cushites, it likely would also have been present in the affiliated Modern South Arabian populations. Instead, the opposite is true: Mehri, as a representative MSA group, are the least admixed Semitic-speaking population. Vyas’ finding therefore supports the view that the Proto-Cushites were not significantly admixed with any Sub-Saharan groups. (*N.B. A few Mehri also settled in the Horn, where their descendants are today represented by the Carab Salaax. This tiny group numbers a few hundred individuals, who dwell in the northeastern Puntland region of Somalia.)

The Makalia skull, which belonged to an early Cushitic settler of the Pastoral Neolithic. Although since lost, Louis Leakey (in the cranium’s original form) and G. P. Rightmire (in its cast form) observed that the specimen shared closest morphological ties with “Mediterranean Caucasoids.”
- Prendergast et al. (2018) analyzed the DNA of various ancient South Cushitic settlers in the Great Lakes region, who belonged to the Pastoral Neolithic (PN) cultural complex. A chronological examination of the mtDNA lineages of these individuals reveals that the earliest PN specimens bear maternal haplogroups that are today most common among Afro-Asiatic speakers rather than clades that are most frequent among Nilotes. In fact, the oldest of the examined Cushitic pastoralists, a 4080-3890 year old individual buried at the Prettejohn Gully site, belongs to the haplogroup K1a. The earliest recorded occurrence of this lineage in Africa is among specimens at Kelif el-Boroud, the aforementioned Neolithic site in Morocco (Fregel et al. note that “K is considered a typical European Neolithic lineage”; see here). Haplogroup K was also borne by the ancient Egyptian Pharaoh Tutankhamun, having been passed down to him from his mother “The Younger Lady”, his grandmother Queen Tiye (“The Elder Lady”), and his great-grandmother Thuya (cf. Gad et al. (2020b)).In terms of Y-DNA, one of the early Cushitic Pastoral Neolithic specimens from Prettejohn Gully was assigned to the E2(xE2b) or E-M75 haplogroup. By contrast, Prendergast et al.’s later Cushitic Pastoral Neolithic individuals overwhelmingly belong to various subclades of the E1b1b haplogroup (72% or 13/18 carriers). The oldest appearance of this lineage and the E clade as a whole in the archaeogenetic record is among Iberomaurusian, Natufian, Neolithic Moroccan and Pre-Pottery Neolithic Levantine specimens. In fact, as with the Afro-Asiatic-speaking populations in North Africa, E1b1b remains today the dominant paternal haplogroup among the Afro-Asiatic-speaking populations in the Horn of Africa. The clade is borne at especially high frequencies by Tigre (100%), Saho (98.9%), Somali (77.6%) and Tigray-Tigrinya (71.9%) individuals (cf. Trombetta et al. (2015), Supplementary Table 7; Sanchez et al. (2005)). This discrepancy between the Y-DNA lineages of the early vs. the later Cushitic Pastoral Neolithic specimens has led Prendergast et al. to speculate that these samples might represent two distinct movements of ancient herders into East Africa, with the later Neolithic pastoralists appearing to be the more demographically impactful. Haplogroup and genome evidence, however, indicates that this was likely not the situation. Hirbo (2011) reports that various Cushitic-admixed Nilote populations in the present-day Great Lakes region actually do bear the E2 haplogroup at significant frequencies (highest among the Tugen at ~61.8%). These groups also have a low incidence of the E1b1b lineage, and they did not receive much maternal gene flow from Cushitic peoples. Yet, parodoxically, Scheinfeldt et al. (2019) observed that these Nilotes harbor the same Cushitic-derived West Eurasian ancestry as the Datog and other such Nilotic populations, who more intensively absorbed E1b1b-carrying Cushitic groups. The Tugen and similar lacustrine populations, therefore, seem to have inherited the E2 clade specifically from their Pastoral Iron Age Nilotic ancestors since a) Prendergast et al. observed that ~67% of the PIA Nilotes carry the E2 haplogroup (2/3 specimens; the other individual bears the E1b1b-M293 clade), and b) the PIA Nilotes, in turn, acquired the E2 and E1b1b paternal lineages and West Eurasian admixture from assimilating Cushitic individuals of the Pastoral Neolithic (we can be confident of this since older Nilotic individuals examined by Yousif and Eltayeb (2009) primarily belong instead to the archaic haplogroup A; see Ancient DNA from Sudan). In sum, this means that the early herders of the Pastoral Neolithic were indeed essentially of the same Cushitic ancestral stock as the later Pastoral Neolithic herders.
-

- Y-DNA haplogroups of modern populations in Ethiopia. Some Oromo, Amhara and Wolayta individuals bear the E1b1b-M293 haplogroup (~17%-28%), which ancient DNA analysis has revealed is the most common paternal lineage of the later Cushitic settlers of the Pastoral Neolithic. However, the M293 subclade is today absent or rare among other Afro-Asiatic-speaking populations in Djibouti, Eritrea and Somalia. Assuming that the Afro-Asiatic speakers in Ethiopia did not obtain E1b1b-M293 through assimilation of Southern Cushitic groups, this suggests that the later Pastoral Neolithic herders arrived from North Africa by way of Ethiopia (Pagani et al. (2015), Supplementary Data).
-

- Y-DNA haplogroups of ancient Cushitic individuals of the Pastoral Neolithic cultural complex. The sampled early Neolithic pastoralist carries the E2 or E-M75 paternal haplogroup, whereas the later Neolithic pastoralists mostly belong to various subclades of the E1b1b haplogroup; primarily M293 (Wang et al. (2020), Supplementary Material).
Only around 1,300 years after the earliest radiocarbon-dated Pastoral Neolithic sample does the first probable evidence of Nilotic influence, in the form of the Y-DNA haplogroup A and the mtDNA clade L4, appear among specimens excavated at the Naishi Rockshelter and Keringet Cave. As the years pass, we can see the Afro-Asiatic-associated mtDNA lineages further giving way to hunter-gatherer and Nilotic-affiliated clades. The Great Lakes would, by the Iron Age, eventually become dominated by its current primary occupants, the Bantus (descended from the Iron Age/IA culture makers) and the Nilotes (descended from the Pastoral Iron Age/PIA people). This gradual population replacement of that region’s early Cushitic pastoralists is also reflected in the osteological record (see G. P. Rightmire’s analysis on The Elongated African fallacy).
-
- Structural genome analysis of the ancient Cushitic pastoralists of East Africa. The earliest of these settlers have been found to derive around 80% of their ancestry from a North African/Middle Eastern ancestral population (red component). At the time of their initial arrival in East Africa, these Cushitic pastoralists presumably would have borne even higher non-African ancestry (~90%) since they would have lacked forager admixture (blue component), which appears to be indigenous to the region (Wang et al. (2020), Supplementary Material).
Wang et al. (2020) analysed additional genomes belonging to the ancient Cushitic pastoralists of East Africa. Like Prendergast et al., the researchers found that the earliest of these Cushitic settlers largely bore North African-related ancestry. They also formulated a three-way admixture model using Levantine Chalcolithic, Dinka and Mota specimens as hypothetical ancestral populations to the ancient Cushitic herders. However, thanks to Wang et al.’s improved and more broad-based dataset (which includes several new Pastoral Neolithic samples from Kenya), we are now able to confirm that the first Cushitic arrivals in the region indeed were not significantly admixed with Nilo-Saharan speakers or other Sub-Saharan groups. Admixture analysis instead shows that the early Cushitic pastoralists had almost identical ancestry proportions as modern Afro-Asiatic-speaking North African populations, deriving around 80% of their genome from a West Eurasian ancestral population (red component). Furthermore, according to both Wang et al.’s admixture analysis and three-way admixture modeling, these earliest Cushitic settlers had just 5%-10% of supposed admixture from Nilo-Saharan peoples (purple). This completely debunks the claim that the Proto-Cushitic speakers, who evolved in the Egypt/Libya/northern Sudan area and subsequently migrated southwards, carried significant ancestry related to Nilotes. The later group of Cushitic Pastoral Neolithic herders were very similar to the earlier Cushitic pastoralists in that they derived ~70% of their ancestry from a North African/Levantine-related ancestral population. At the time of their first settlement in the region, these ancient Cushitic peoples would have borne even higher non-African ancestry (~90%). We know this because they were also found to carry some East Africa-specific forager admixture (blue component), which they must have picked up locally after their arrival (cf. Supplementary Material; Tables S1 to S10).
Moreover, Wang et al. examined two of the ancient Cushitic settlers for phenotype alleles (a 2300 year old sample from Hyrax Hill and a 1500 year old sample from Molo Cave, both in Kenya). The scientists reported that both of the specimens bore the derived allele of the SLC24A5 gene, which is associated with lighter skin pigmentation. This suggests that the individuals had a light brown complexion (cf. Table S7).
(*N.B. Wang et al. also compared the ancient Cushitic pastoralists’ genomes with those of various modern samples, including individuals from the Cushitic-speaking Afar, Agaw, Somali and Oromo groups (cf. Supplementary Material). The researchers did not include any Abyssinian/Habesha/Ethiosemitic-speaking samples in their admixture analysis, presumably because Abyssinians are not the best representatives of the early Cushites since they interbred with Semitic and Omotic peoples (Wang et al. genomically model the Agaw sample as having 5.6% more Levant Chalcolithic ancestry than the Amhara test group due to the Amhara cohort’s higher Omotic/Mota-related admixture; see Tables S1 to S10). In the admixture analysis, the Afar and Agaw samples had virtually the same estimated ancestry proportions as the later Pastoral Neolithic samples. However, the Oromo and Somali samples had slightly less of the North African-related component. This is probably because the Ajuran and Garre Somali clans in Kenya have intermarried with Borana Oromos (cf. Helland (1980)), and Borana Oromos have some Omotic admixture (the Somali sample in Wang et al. (2020) was taken from Prendergast et al. (2018), who used the Simons Genome Diversity Project’s Somali sample; this SGDP sample was collected in Kenya by George Ayodo). Future analyses should be sure to incorporate Oromo samples from central Ethiopia, which are less affected by Omotic admixture. They should also include ethnic Somali samples from the overlooked Puntland and Somaliland regions in northern Somalia.)
-

-

- Facial skin color among Somalis. Most individuals have lighter tones (chiari), which correlate with tones 25 to 29 on the Von Luschan Chromatic Scale. A minority of individuals possess darker tones (scuri), which correspond with the Von Luschan Chromatic Scale’s 30 to 33 tones. We now know from ancient DNA analysis that the lighter tones were inherited from the ancient Cushites, who had a relatively fair complexion (Puccioni (1931)).
-

- Skin tones on Von Luschan’s Chromatic Scale.




Young ethnic Somali men in the southern Jubaland region of Somalia (top), young ethnic Somali women in the northeastern Puntland region of Somalia (center top), young ethnic Somali women in the northwestern Somaliland region of Somalia (center bottom), and young Bilen/northern Agaw women in Eritrea (bottom). Genetic analysis has shown that around 60% of Cushitic, Ethiosemitic and North Omotic-speaking individuals in the Horn of Africa carry the derived allele of the SLC24A5 gene. This lighter skin conferring variant has been measured to have an effect on pigmentation of 7.6–11.4 average melanin units (cf. (Beleza et al. (2013)).
- Anthropological examination of the physical remains of the Pan-Grave culture bearers (referred to by the ancient Egyptians as Medjay) indicates that they were a community of heterogeneous origins. Strouhal and Jungwirth (1979) analysed skeletons excavated from five Pan-Grave cemeteries at Sayala, Nubia, which date from the late Roman-early Byzantine period. Of these burial complexes (referred to as CI-III, N and A), the scientists note that the CI-III and N cemeteries predominantly contained the graves of individuals of “Europoid” (“Caucasoid”) physical type, whereas the A cemetery primarily contained the graves of individuals of mixed “Europoid-Negroid” physical type:
The general morphology of the skeletal remains, expressed in classical racial terms, showed a somewhat different picture. The results of the physical anthropological study pointed to the prevalence of the Europoid type (84.5%) in the burial complexes CI-III as well as in the N cemetery, with a small share of mixed (11.4%) and Negroid (4.1%) types. On the other hand, people buried in the A cemetery were mostly of an intermediate morphology (57.1%) with more Europoids (33.3%) than Negroids (9.6%).
Of the few “Negroid” individuals living amongst the mostly “Europoid” Pan-Grave culture bearers, research has found that the former showed archaic characteristics, possessing robust bones with strong muscular attachments as well as distinctly “Negroid” features and woolly hair (Strouhal and Jungwirth (1984); Säve-Söderbergh (1989); Friedman (2001); Brunton (1937)). By contrast, individuals belonging to the Afro-Asiatic-speaking Kerma, C-Group and ancient Egyptian cultures generally had lithe builds, with “Caucasoid” features and non-woolly hair (for specifics on the physiognomy of the Kerma and C-Group culture bearers, see The Elongated African fallacy). Furthermore, Strouhal and Jungwirth (1979) compared the discrete/non-metric osteological traits of the mainly “Europoid” individuals from the three CI-III Pan-Grave cemeteries at Sayala against those of the Pan-Grave specimens from the N cemetery and A cemetery, as well as against those of various ancient Egyptian series (Badari, early Naqada, late Naqada, Hierakonpolis, Abydos, Tarkhan, Sedment, Qurna, Qau, Giza and Hawara), a C-Group Nubian sample (thought to represent early Berber-speaking inhabitants of the Nile Valley), a modern East African “Negroes” sample (Rwanda and Burundi), and a modern South African “Negroes” sample (Zulu, Xosa, Sotho and Venda). The researchers observed that the CI-III specimens were morphologically distinct from all of these comparative samples, a fact which they suggest indicates that the Pan-Grave people of the CI-III burial complex likely represent the Blemmyes of the Eastern Desert. According to Winstedt (1909), an early Coptic inscription refers to the Blemmyes as the Bougaioi. During the later Aksumite era, this name is rendered as Beya in an Adulis text — a clear allusion to the Cushitic-speaking Beja people. This morphological dichotomy between the Pan-Grave culture’s “Europoid” Cushitic majority and its assimilated “Negroid” Nilotic minority is also reflected in ancient artwork. To this end, a 12th Dynasty Nile Valley mural depicts a “Caucasoid” Cushitic Beja figure, whereas a painted ox skull from the same epoch portrays a “Negroid” Nilotic figure.
-

- Ancient Cushitic man depicted on a Twelfth Dynasty Egyptian mural (Seligmann (1913)). Note the figure’s lithe build, fine features, facial orthognathism and loose wavy hair. Skeletal analysis of ancient individuals excavated from four Pan-Grave cemeteries at Sayala (burial complexes referred to as CI-III and N) indicates that most of these specimens had a similarly “Europoid” or “Caucasoid” physiognomy (84.5%) (Strouhal and Jungwirth (1979)).
-

- A Bisharin Beja man, bearing a striking resemblance to the ancient Cushitic mural figure.

A Somali man, retaining the fine features, facial orthognathism and soft-textured hair of his Cushitic ancestors.
-

- Ptolemaic era portrait of an Egyptian man. The figure’s very curly hair is mostly a legacy of admixture with the Hyksos, Semitic speakers who entered the Nile Valley during the Bronze Age. Forensic analysis of earlier Egyptian individuals dating from the predynastic period indicates that most of the examined specimens had soft-textured wavy or straight hair, with some individuals also having soft-textured curly hair (cf. Elliot Smith (1911)).
-

- Egyptian men, retaining the soft-textured curly hair of their Ptolemaic-era Egyptian ancestors.
-

- Nilotic men from areas south of the fourth cataract, shown on a New Kingdom Egyptian frieze with characteristic “Negroid” features, facial prognathism and kinky hair. Skeletal analysis of ancient individuals exhumed from the A cemetery at the Pan-Grave site of Sayala indicates that 9.6% of these specimens had a similarly “Negroid” physiognomy (Strouhal and Jungwirth (1979)).
-

- Nilotic “Negroid” man depicted on a painted ox skull from a Pan-Grave burial at Mostagedda, Upper Egypt.
-
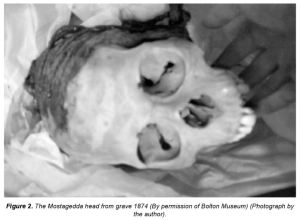
- Skull of an individual excavated from a burial associated with the Pan-Grave culture at Mostagedda. The specimen has an intermediate “Europoid-Negroid” physiognomy. Skeletal analysis of Pan-Grave culture bearers from the A cemetery at Sayala indicates that most individuals at this site had a similarly mixed “Europoid-Negroid” physical type (57.1%). However, this intermediate physiognomy was uncommon among the predominantly “Europoid” individuals buried at the four CI-III and N Pan-Grave cemeteries in Sayala (11.4%) (Strouhal and Jungwirth (1979)).
-

- Old Kingdom statue of an ancient Egyptian man of mixed Egyptian and “Negro” origin. Craniometric analysis of modern “Black Egyptians” indicates that they do not cluster with ancient Egyptians like modern Afro-Asiatic speakers from North Africa and the Horn of Africa do. Rather, due to their mixed background, “Black Egyptians” are morphologically intermediate between ancient Egyptians and Nilo-Saharan/Niger-Congo peoples (cf. Billy (1981b)).
-

The alveolar index (gnathic index) among global populations, which measures levels of prognathism (facial projection). The Somali (94), ancient Kerma (96.7), and early and modern North African samples have a similarly low alveolar/gnathic index as the ancient and modern European samples (Hanihara (2000)).
Interbreeding between European individuals and Nilotic or Bantu individuals will not produce any of the three main phenotypes that are found today among the northern Cushitic and Ethiosemitic-speaking populations in the Horn of Africa (see discussion below on the mariin, cad and yusuur phenotypes). This obvious fact is frequently overlooked by geneticists when constructing their statistical admixture models, probably because they often are not trained in physical anthropology.
Rather than anthropometrically clustering in between Europeans and Nilotic/Bantu individuals like actual biracial persons do, the Afro-Asiatic speakers in the Horn region group with other Afro-Asiatic speakers in North Africa and the Arabian peninsula (cf. Billy (1988); Leguebe (1981)). Instead of having nasal indices between those of Europeans and Nilotes/Bantus like actual biracial persons have, the nasal indices of the northern Cushitic and Ethiosemitic-speaking populations also fit comfortably within the “Caucasoid” range (Olapido et al. (2011)). Likewise, Gedda (1961) indicates that “the nasal breadth of the American Negro (40.9) is much less than in African Negroes (42.4-45.5), that for White Americans being 34.9 and for Somali only 34.7.” Skeletal analysis of ancient Cushitic specimens by Louis Leakey observed similarly leptorrhine (narrow) nasal indices (see The Elongated African fallacy). This was, therefore, the original nasal form of the Cushitic peoples.

Oromo women, possessing the “Caucasoid” physiognomy and soft-textured hair characteristic of “pure” Cushitic people.
— Somali men with the two most common hair forms distributed among ethnic Somali males (over 80% in non-Sab clans). These consist of wavy strands and frizzy curls, both of soft-texture (jileec) (Coon (1939); Puccioni (1937)).
Hair form provides a similar indication, for Coon (1939) notes that approximately 86% of the Somalis he examined had soft-textured curly, wavy or straight hair (also see Charpin and Georget (1977) chart above). The autonym jileec (“soft-haired”), which ethnic Somalis traditionally use to describe themselves, suggests that this was the original hair form of the Cushitic peoples (Bernus (2000)). Indeed, the medieval Portuguese missionairies João dos Santos and Jerónimo Lobo state that the Maracatos (Katwa or Garre Somali) — notorious slave traders who castrated their slaves (cf. Steiner (1999)) — that they encountered in southern Somalia were tall, swarthy people with fine-features and lank hair (cabello corredio). We also know from forensic analysis that the early Cushitic speakers of Kerma in Sudan almost unanimously possessed non-kinky hair texture (Lepsius (1915)). Contrarily, many if not most European-Nilotic/Bantu biracial individuals have kinky hair texture like their Nilotic/Bantu parent (this often does not apply to New World biracial persons, who, due to the extra European admixture and Native American admixture that is already present in their African American/Afro-Caribbean parent, may have soft-textured hair).
-
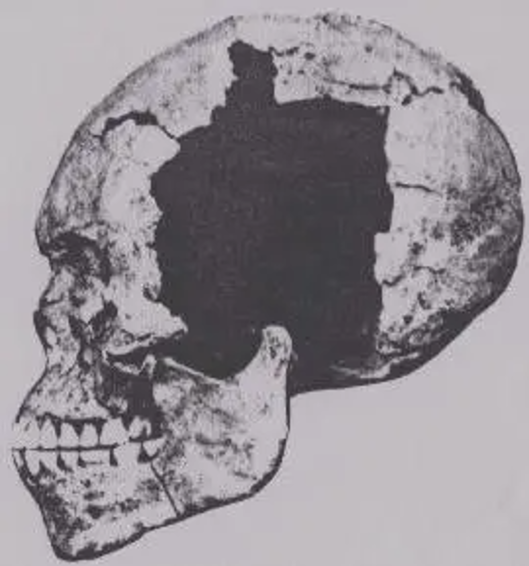
- A Mesolithic period “Negroid” skull excavated in Wadi Halfa, Sudan. The specimen is believed to be ancestral to the local Nilotic populations, as it closely resembles the crania of modern Nilo-Saharan/Niger-Congo peoples. Note the marked prognathism (facial projection). This is a common trait of Nilotes and other African “Negroes,” who have a prognathous average of 104.4 on the gnathic index (alveolar index) (cf. Lockyer (1910)).
-

- A Nilotic Dinka man, retaining the characteristic facial prognathism of his Nilo-Saharan/Niger-Congo (“Negro”) ancestors.

Ancient Egyptian engravings of an early Cushitic individual (left) and an early Nilotic individual (right). The contrast in physiognomy between the two is evident, pointing to different ancestral origins. The Cushitic individual is fine-featured and lean, and has soft-textured wavy hair, medium-sized and non-everted lips, and an orthognathous profile. Conversely, the Nilotic individual has broad and flat facial features, plaited kinky hair, large everted lips, and is markedly prognathous.

A Hadendoa Beja man preserving in all essential aspects the physiognomy of the engraved ancient Cushitic figure above.
Furthermore, instead of sharing at least some of the prognathism (facial projection) that characterizes Nilo-Saharan/Niger-Congo groups, the Afro-Asiatic speakers in Northeast Africa are among the most orthognathous populations in the world. Hanihara (2000) thus observed that the alveolar index (gnathic index), which measures prognathism levels, is particularly low among his Somali sample (94); the same goes for the ancient and modern North African samples, including the Kerma cohort (96.7). Leigh (1934) similarly indicates that “the ancient Egyptians have a low gnathic index, my average for sixty-nine males being 95.2, and for seventy-one females, 95.6.” By contrast, Wellcome Tropical Research Laboratories (1908) report that the Nuer and Bari Nilotes of the Sudan area have a “very considerable” gnathic index of 106.6 and 109, respectively. This is an even greater level of prognathism than that measured among the Niger-Congo-speaking samples. In fact, some of the highest gnathic values ever have been recorded among the Nilotic peoples. Lockyer (1910) reports on the skull of a Nilotic woman (dated to c. 2nd-4th centuries CE), which had an extreme gnathic index of 123.3, almost 19 points higher than the 104.4 prognathous average of African “Negroes.” (*N.B. With regard to the alveolar index or gnathic index, craniofacial profiles that are orthognathous=below 98, mesognathous=98.1-103, and prognathous=above 103.)
-
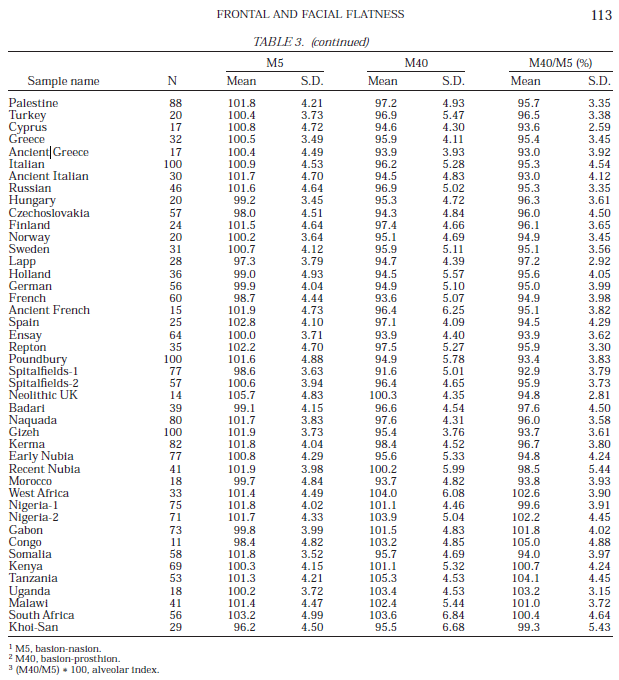





-

- Sandra Banks, former Miss South Sudan (European father & Nilotic mother). Banks resembles most biracial individuals born to a European parent and a Nilotic or Bantu parent in that her physiognomy is intermediate between the two, though favoring her Nilotic side overall.
-

- A Nilotic Dinka woman, largely retaining the facial features, hair texture and skin complexion of her Nilote ancestors despite some Cushitic admixture.
-

- Chiara de Blasio (European father & African American mother). De Blasio resembles most biracial individuals born to a European parent and an African American/Afro-Caribbean parent in that her physiognomy is intermediate between the two, though favoring her European side overall (except for hair texture). This is due to the extra European admixture that is already present in her African American parent.
-

- Chirlane McCray, mother of Chiara de Blasio (African American). Like most African American individuals (particularly in the southern USA), McCray has largely retained the physiognomy of her Niger-Congo ancestors despite having significant European and Native American admixture.
-

- Alexander Yusuf, architect (Somali father & British mother). Individuals with Cushitic and European parentage often look almost completely European. This is because they are the offspring of a European parent and a predominantly West Eurasian parent (Cushitic side). Ancient DNA analysis has shown that the earliest Cushitic peoples were of North African ancestral stock (~80% non-African ancestry), and their modern descendants in the Horn of Africa have comparable affinities (almost 70% non-African ancestry, after correcting for linkage disequilibrium bias).
-

- Ella Thomas, actress (European father & Tigrinya mother). Other individuals with Cushitic and European parentage can instead pass for regular Horn Africans with the mariin (“red brown”) phenotype. This usually occurs when their Horn African parent has more pronounced, visible Ari or other Sub-Saharan admixture (i.e. the yusuur phenotype). The West Eurasian genes from the person’s European side, therefore, have the effect of counterbalancing that extra Sub-Saharan admixture. This then brings the individual’s physiognomy closer to that of “purer” Cushitic peoples.
-

- A half Somali, half European woman. The girl is of ordinary European appearance, which is the norm for individuals born to a European parent and a Cushitic parent with the cad (“olive-skinned”) phenotype. This physiognomy is also rather common among individuals born to a European parent and a Cushitic parent with the mariin (“red-brown”) phenotype, though here the offspring tend to be a bit swarthier.
-

- A half Eritrean, half Serbian woman. Individuals born to a Cushitic parent and a European parent often look strongly Mediterranean or Sardinian-like, with small, delicate facial features, fine straight/wavy hair, and brunette hair and eye coloring. According to Molinaro et al. (2019) and Aneli et al. (2021), among other researchers, this similitude is an atavistic trait, harkening back to the “Pan-Mediterranean” ancestral population from whom the early Cushites descend.
- As pointed out earlier in the essay, craniometric analysis does not support the idea that the modern Afro-Asiatic-speaking populations in the Horn of Africa experienced significant gene flow from Nilotic/Bantu/Khoisan communities, to the point where they would cluster with racially mixed African groups such as Black Egyptians (cf. Billy (1981b)). The Cushitic and Ethiosemitic speakers instead show closest affinities with ethnic Egyptians and other Afro-Asiatic-speaking populations in North Africa and the Middle East, in addition to Europeans and other peoples with a generally “Caucasoid” craniofacial pattern (Brace (1993); Spradley (2006)). Billy (1977) thus notes that craniometrically “the modern Hamite Ethiopian series (Tigre) and even Somali-Galla remain virtually exempt of any black contribution. The Ethiopian plateaus must therefore have remained far from the currents of black immigration which took the natural route of the Nile, whether during the dynastic period or later and more intensively, from the beginning of our era on.” This is consistent with the observation above that there is a lot of extra non-African ancestry in the genomes of the Afro-Asiatic speakers, which is not being properly detected.
-
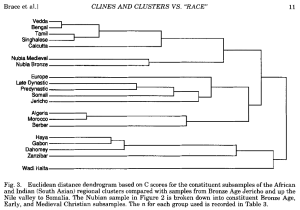
- Craniometric affinities of ancient and modern populations worldwide. Somalis cluster with the North African, ancient Egyptian, Bronze age Jericho and European samples, whereas the Zanzibaris/Swahilis (despite having some recent west Eurasian admixture) group with the Bantu and West African samples (Brace (1993)).
-

- Craniometric affinities of various populations in Africa and the New World. Somalis and Egyptians cluster together, while African-Americans (despite having recent European and Native American admixture) group with the West African samples (Spradley (2006)).
-

- Craniometric affinities of ancient and modern populations in Africa. The Tigre and lumped Somali-Galla samples cluster with the ancient Egyptians and early Afro-Asiatic-speaking populations of Nubia, whereas the Black Egyptians/Noirs Egyptiens (despite having recent Egyptian admixture) group with the ancient Nilotic and modern Bantu samples (Billy (1981b)).
-

Structural genome analysis of Afro-Asiatic-speaking and Nubian populations in Northeast Africa. At K=7, Cushitic ancestry (yellow component) is maximized among the Somali North sample (Ethiopian Somali). The Somali South sample (Sab Somali) is most similar to the Somali North sample, but differs from it by having greater Sub-Saharan admixture (Nilo-Saharan/orange & Niger-Congo/purple components) (Vicente et al. (2019)). This is consistent with anthropometric (Billy (1988)) and craniometric (Rightmire (1976)) analyses of these groups. (*N.B. Vicente et al. (2019) indicate in their Table S1 that their “Somali North” sample was taken from Gurdasani et al. (2015). As explained above, Gurdasani et al. stipulate that “although these samples were collected from Ethiopia, many of these individuals are from Somalia (Mogadishu).” Hence, Vicente et al. (2019)’s “Somali North” sample is in actuality primarily a Mogadishu Somali or South-Central Somali sample (i.e., Hawiye Somali). Furthermore, Vicente et al. state in their Table S1 that their “Somali South” sample was borrowed from Triska et al. (2015). Triska et al., in turn, note in their own Table S1 that their Somali sample consists of 11 individuals who speak an Afro-Asiatic language, practice a sedentarian subsistence system, and originate from the geographical coordinates of 42.63 longitude and 2.00 latitude. GPS-coordinates.org specifies that these geographical coordinates correspond to the town of Diinsoor, located in the southwestern Bay region of Somalia. This is an area almost exclusively inhabited by the Rahanweyn or Sab Somali agropastoralists. I. M. Lewis writes that “the Sab are a conglomerate people, an amalgam of many different Somali groups with Galla and negroid elements,” thus explaining the excess Sub-Saharan African admixture in this “Somali South”/Sab cohort.)
The anthropometric data measured by Puccioni (1931) and Coon (1939) suggests that the populations responsible for the African admixture component in the Horn of Africa were generally of short stature like the Mota, Gogoshiis Qabe and Buur Heybe hunter-gatherers rather than of tall stature like Nilotes. This is indicated by the fact that the most African-admixed groups among the Afro-Asiatic speakers in both Ethiopia and Somalia also happen to be the shortest on average (viz. Sidama and Sab), although they are still taller than the local foragers (viz. Chabo, Sabue, Eyle, Boni, Watta and Ribi; Puccioni reports an average height of 165.15 cm/1651.5 mm for the Ribi). Conversely, Puccioni observed that there was a positive correlation between high stature, lighter skin pigmentation and non-woolly hair texture in his cross-analysis. Patenostre (1922) likewise found that his Somali sample had a crural index or tibio-femoral index (which estimates leg limb proportions) of 83.2, similar to the 83.4 average of the Berber and Arabic-speaking populations of North Africa. Both Afro-Asiatic-speaking groups were also distant from the “Negro” sample, which had a crural index of 86.8. Moreover, genome analysis of the Early Pastoral Neolithic specimens of Kenya, who are the oldest Cushitic settlers in eastern Africa to be genetically analysed, indicates that they bore little Nilo-Saharan-related ancestry (8.5% on average). Since the rest of their Sub-Saharan admixture consists of a minor East African hunter-gatherer element (~14.7%), which is native to the region, this implies that these first Cushitic arrivals in eastern Africa would have borne almost exclusively non-African ancestry at the time of their first settlement in the area (~91.5%). Taken together, this supports Carleton Coon’s theory that the moderately tall height of modern Cushitic speakers was directly inherited from their “Caucasoid” Cushitic ancestors as opposed to acquired by mixing with Nilotes (as Coon phrased it, “there can be no doubt that the tall stature of the Gallas, Somalis, and Agaus is an old Hamitic trait[…] The bodily build of the East African Hamites is typically Mediterranean in the ratio of arms, legs, and trunk”). (See Genetic affinities of the Cushitic, Ethiosemitic and North Omotic-speaking populations of the Horn of Africa for details.)

Crural index (tibiofemoral index) of various populations in Africa and Europe (Patenostre (1922)).
-

- Ribi man. Although now admixed with their Cushitic neighbors, the diminutive Ribi foragers of southern Somalia are thought to be direct descendants of the native hunter-gatherers of the eastern Horn.
-

- Acculturated man of Ribi hunter-gatherer origin. Such non-Cushitic individuals are known in the Horn as sheegad (“pretenders”) due to the fact that they are attached to the Afro-Asiatic-speaking groups on a traditionally subordinate, adoptive/client basis. The man’s close resemblance to the Ribi and other indigenous foragers thus stems from shared ancestral roots.
-

- Sandawe women, showing little Cushitic physical and cultural influence (Bisa Sandawe). In the Great Lakes region, Sandawe individuals of “purer” hunter-gatherer type are centered in the Bisa, the ‘original’ division of the Sandawe community. The Bisa Sandawe also generally still practice the foraging lifestyle of their hunter-gatherer ancestors.
-

- Sandawe man with substantial Cushitic admixture (Tehla Sandawe). Many early Cushitic pastoralists were absorbed into the Tehla, the herder division of the Sandawe. Consequently, Sandawe individuals today, like the Boni, carry moderate levels of West Eurasian ancestry (~30%; cf. Scheinfeldt et al. (2019)). A significant minority of Sandawe also bear the Afro-Asiatic-linked E1b1b paternal haplogroup (24.3%). This altogether indicates that the Sandawe’s hunter-gatherer ancestors had extensive contact with ancient Cushitic settlers (Hirbo (2011)).
-


-
- Linguistic analysis of the Afro-Asiatic languages spoken today in the Horn of Africa has not identified any Nilo-Saharan substratum in these idioms. For instance, Amharic and Tigrinya instead have a Cushitic substrate because their speakers originally spoke Agaw languages. A linguistic substratum, of course, isn’t a prerequisite for population admixture. Nonetheless, such features are often retained when one speech community shifts to speaking another language. The conspicuous lack of a Nilo-Saharan substratum in the region’s main Afro-Asiatic languages is therefore congruent with the notion that gene flow between the two speech communities was largely unidirectional. That is, it was the early Cushitic speakers who primarily impacted the Nilo-Saharan speakers rather than vice versa. (*N.B. The above principle does not apply to languages such as Daasanach, which does have a Nilo-Saharan substratum since its current speakers originally spoke a Nilo-Saharan language. Consequently, Poloni et al. (2009) and López et al. (2021) observed that their sampled Daasanach (Dasanech) individuals are genetically more similar to Nilotic populations inhabiting the Great Lakes region than to other Afro-Asiatic-speaking groups in the Horn of Africa.)
-
-
-

Tuareg Berber nobles of the Kel Tamasheq confederation.
Among the northern Cushitic groups in the Horn region, three main phenotypes are distinguished: mariin (“red-brown”), cad (“light”) and yusuur (“ebony”). The mariin phenotype is the most widespread, and is characterized by tall stature, leptorrhine facial features, copper-brown skin complexion, soft-textured curly, wavy or straight hair, medium-sized lips with no eversion, dolichocephaly, orthognathism and oftentimes slightly oblique eye folds (cf. Puccioni (1931); Gallo (1979)).
-

- Beja men with the amrani/mariin phenotype.
-

- A Somali man with the amrani/mariin phenotype.
-

- A northern Sudanese man with the amrani/mariin phenotype.
-

- An Egyptian man with the amrani/mariin phenotype.
-

- Jordanian Arab bedouin men with the amrani/mariin phenotype (Ammarin tribe).
-

- An Iraqi Arab bedouin man with the amrani/mariin phenotype.
-

- Yemeni men with the amrani/mariin phenotype (Bagefer tribe).
-

- A Yemeni man with the amrani/mariin phenotype (Marib governorate). Genome analysis has found that, after the Mahra or Mehri, Yemeni individuals from Marib have the most Natufian ancestry of all modern populations.
-

- Algerian singer Cheb Khaled with the amrani/mariin phenotype.
-

- Somali politician Abdirahman Jama Barre with the amrani/mariin phenotype (Darod clan).
-

- Bedouin Arab man with the amrani/mariin phenotype (Negev Desert, Israel).
-

- Omani Sultan Qaboos bin Said with the amrani/mariin phenotype.
-

- A Somali man from Djibouti with the amrani/mariin phenotype (Dir clan).
-

- Sheikh Hasan Aly Mustafa of the Ababda Beja with the amrani/mariin phenotype.
-

- An Egyptian man with the amrani/mariin phenotype (Kharga Oasis).
-

- A Nubian man with the amrani/mariin phenotype.
-

- A Shluh Berber man with the amrani/mariin phenotype.
-

- A Tuareg Berber man with the amrani/mariin phenotype.
-

- Al-Saud royal family of Saudi Arabia with the amrani/mariin phenotype. Before hitting it big with oil wealth and marrying the fairest brides available to them, the Al-Sauds had a reddish-brown complexion like many other Bedouin Arabs.
-

- Maghrebi Sultan Moulay Hafid with the amrani/mariin phenotype.
-

- Saudi Prince Saud Al Kabeer bin Abdulaziz Al Saud (center) with the amrani/mariin phenotype.
-

- Somali Sultan Yusuf Ali Kenadid (center) with the amrani/mariin phenotype (Darod clan).
-

- Somali leader Muhammad Abdullah Hassan (the “Mad Mullah”) with the amrani/mariin phenotype (Darod clan).
-

- A Yemeni Arab camel breeder (Semitic).
-

- Yemeni Sultan al-Fadl III ibn ‘Ali al-‘Abdali with the amrani/mariin phenotype (Lahij, Yemen).
-

- Somali Sultan Ali Yusuf Kenadid with the amrani/mariin phenotype (Darod clan).
-

- Toubou leader Kossey Sanda with the amrani/mariin phenotype.
-

- Emirati Arab Sheikh Zayed bin Khalifa Al Nahyan (“Zayed the First”) with the amrani/mariin phenotype.
-

- A Socotri man with the amrani/mariin phenotype.
-

- An Assyrian man with the amrani/mariin phenotype. Note the close resemblance to the Socotri man. This shared likeness highlights the fact that, despite mixing significantly with light-skinned outsiders from the Caucasus/Iranian plateau, many modern Assyrians have retained the swarthy complexion of their Semitic ancestors.
-

- A Nubian man with the amrani/mariin phenotype.
-

- An Afar man with the amrani/mariin phenotype.
-

- An Al-Nasir Sudanese “Arab” man with the amrani/mariin phenotype.
-

- A Syrian Arab bedouin man with the amrani/mariin phenotype.
-

- Jordanian Arab bedouin men with the amrani/mariin phenotype.
-

- Rendille comedian Meleyo Saigor with the amrani/mariin phenotype.
-

- An Arab Chief with the amrani/mariin phenotype (Dhufar, Oman).
-

- Somali scholar Osman Yusuf Kenadid with the amrani/mariin phenotype.
-

- A Beja man with the amrani/mariin phenotype.
-

- An Egyptian man with the amrani/mariin phenotype.
-

- A Rashaida Arab man with the amrani/mariin phenotype.
-

- Saho scholar Istaz Ahmed with the amrani/mariin phenotype.
-

- Jordanian Arab bedouin men with the amrani/mariin phenotype.
-

- An Arab Amir with the amrani/mariin phenotype (Bani Bu Ali tribe, the Bliulaie of Ptolemy’s Geographia).
-

- Ancient Egyptian figures depicted with the standard amrani/mariin (right) and beidan/cad (left) phenotypes.
-

- Ancient Egyptian mural of the nobleman Sennefer and his wife Merit (18th Dynasty). Sennefer is depicted with the amrani/mariin phenotype.
Other examples:
-

- A Saudi Arab bedouin man with the amrani/mariin phenotype.
-

- An Afar man with the amrani/mariin phenotype.
-

- An Abyssinian man with the amrani/mariin phenotype.
-

- A Mozabite Berber man with the amrani/mariin phenotype.

Muna Khalif (Tuni, Rahanweyn Somali). Although cad individuals in Somalia are concentrated in the northern Puntland and Somaliland regions, a number of persons with this phenotype, such as fashion designer Muna Khalif, can also be found in southern areas.
The cad phenotype is essentially the same as the mariin one, with the exception that cad individuals have a distinctive olive to light brown skin tone. This physiognomy can be found throughout the Horn, but is largely concentrated in northern Somalia, eastern Ethiopia, and parts of Djibouti and Eritrea. Cushitic individuals with the mariin and cad phenotypes, the “pure” Cushitic physiognomies, have traditionally been hailed for their good looks. They were often described in the literature as beautiful, handsome or prepossessing (e.g. the explorer Thomas Heazle Parke writes: “The Somali’s type of feature is Asiatic[…] These Somalis are also very agile in their movements–compared with the Negroes[…] Their skin is of a coppery tint; the natural expression of the face is strikingly bright–with quick, dark, and very mobile eyes–and, altogether, their appearance is prepossessing”). This custom dates back to Herodotus, who asserted that the ancient Macrobians (thought to be an early Cushitic people) were the “handsomest race of men in the world.”
The yusuur phenotype also resembles the mariin one, except that individuals with this physiognomy frequently show traces of admixture with the aboriginal hunter-gatherers of the Horn in that they have a blackish-brown skin tone and sometimes shorter height and frizzier hair. This phenotype occurs across the region, but is most common in southern Somalia, particularly among the Sab (Puccioni (1931); Coon (1939)).
-

- Somali man with the mariin phenotype. This “pure” physiognomy is a reddish-brown version of the cad morphology of the early Cushites. As such, both phenotypes are well-represented on ancient Nile Valley murals and frescoes. The mariin phenotype is still widespread among the modern Afro-Asiatic-speaking populations in Northeast Africa and the southern Maghreb, as well as among many Nubians.
-

- Afar man with the yusuur phenotype. This “mixed” physiognomy is found across the Horn, but is most common in southern Somalia, parts of Djibouti, Ethiopia and Eritrea. It is related to the mariin phenotype, having evolved in the Horn through contact between the region’s early Cushitic settlers (of cad and mariin phenotypes) and the indigenous Ribi hunter-gatherers.
-

- Somali politician Hassan Mohamed Hussein “Mungaab” (Hawiye clan), with an example of the mariin (“red-brown”) phenotype among individuals in south-central Somalia.
-

- Somali politician Mohamed Nur “Tarzan” (Hawiye clan), with an example of the yusuur (“ebony”) phenotype among individuals from the same clan in south-central Somalia.
-

- Amhara Abyssinian leader Ras Nasibu Emmanual with the mariin phenotype.
-

- Amhara Abyssinian Emperor Menelik II with the yusuur phenotype.
According to Coon, the mariin and cad phenotypes found especially among Somalis closely resemble the amrani and Ihaggaren (beidan) phenotypes, respectively, which are characteristic of the Tuareg Berber nobles in the southern Maghreb.

Serological analysis of the Tuareg Berbers, the Ikelan (the slave class within Tuareg society), and other populations of the Sahara and Sahel. The Tuareg individuals cluster with the Arabic-speaking Maghrebi samples. The Ikelan group instead with the Niger-Congo-speaking Serer, Wolof and Toucouleur (Haalpulaar), and more remotely with the Fulani (Peul) of Senegal and the Haratin (Froment (1999)).
Note that the amrani/mariin and beidan/cad phenotypes are not characteristic of the Tuareg’s slave class, known as Ikelan, whose members instead have a “Negroid” physiognomy (see Coon’s discussion below). Genetic analysis has confirmed that the Ikelan are of West African origin since they carry the E1b1a paternal haplogroup, which today is the modal clade among Niger-Congo speakers. The Tuareg nobles, on the other hand, are of Berber origin. They thus primarily belong to the E1b1b lineage, which is typical of the Afro-Asiatic-speaking populations in North Africa and the Horn of Africa (cf. Ottoni et al. (2011)):
The Tuareg nobles are tall men, with mean statures running tribally from 172 cm. to 178 cm.; about 174 cm. would be their total mean. They are lean, long-armed, and long-legged, with narrow shoulders, narrow hips, and chests which are narrow in an antero-posterior direction; their hands and feet are long and very narrow, their fingers long and thin. The very fine wrists and ankles which we have observed among the Somalis are also present here. The addition of Negro blood to this Tuareg bodily type broadens the shoulders, shortens the legs, and makes the hands and feet wider and larger. The Tuareg relative sitting height mean of 49, indicating that the sitting height is less than half the stature, serves to illustrate the extremely linear constitutional type of this people. The heads of the Tuareg are dolichocephalic and large; tribal means in the cephalic index vary between 72 and 75, but 73 is the central point of the whole. No brachycephals are found among the white nobles, although they occasionally appear among negroids of other classes. […]
The Tuareg in their pure form belong to a specialized Mediterranean sub-type, the creation of which is partly a matter of isolation and selection under extreme environmental stimuli. They resemble the East African Hamites very closely, and especially the whiter element among the Somali, but in their extreme stature and great head size they seem closer than most other living Mediterraneans to the pre-Neolithic East African men.[…]
the unexposed skin of the non-negroid nobles and Imrad is seen to be a brunet-white, without brownish tinge; the mixed bloods, however, who are predominantly Tuareg and only in a minor degree negroid, assume a constant and characteristic dark brown color, known in North Africa as amrani, and comparable to the characteristic hue of the Somalis.
-

- Distribution of facial and body hair among Somalis. Most individuals have sparse facial and body hair, with a thin mustache. This is similar to the ancient Egyptians, relatives of the early Cushites. On average, less than 10% of persons have abundant facial and body hair (as common among Semitic individuals) or missing facial and body hair (as common among Niger-Congo/Nilo-Saharan/Khoisan individuals) (Puccioni (1931)).
-

- Distribution of lip forms among Somalis. Most individuals have medium-sized lips, with no eversion (Puccioni (1931)).
-

- Nasal profile among Somalis. Most individuals have a straight nasal profile (rettilineo) (Puccioni (1931)).
-

- Forehead shape among Somalis. Most individuals have a straight (diritta) or slightly receding (poco sfuggente) forehead, with few possessing a very receding (molto sfuggente) or prominent (prominente) forehead shape (Puccioni (1931)).
-

- Average skin pigmentation reflectance among Amhara Abyssinians in Ethiopia. The values correlate with the mariin/amrani phenotype, which indicates that most of the individuals have a copper-brown complexion (Harrison et al. (1969)).
-

- Skin color reflectance values among Jews and other peoples of Middle Eastern origin are intermediate between those of Europeans and most Africans. These values often correspond with the light brown to olive complexion of North African and Cushitic individuals with a beidan/cad phenotype (Lourie et al. (1973)). They are also consistent with Jablonski and Chaplin (2000), who indicate that Israelis have similar skin pigmentation reflectance values as Lebanese and other individuals of the northern Middle East.
The persistence of the mariin and cad phenotypes today in the Horn of Africa further supports the idea that the earliest Cushitic settlers in the region were not significantly admixed with Nilotic populations, for, if they had been, their modern descendants would not continue to resemble the amrani and beidan Tuareg Berber nobles so closely.
-

- Somali diplomat Halima Ahmed with the amrani/mariin phenotype.
-

- Algerian “Arab” socialite Hayat Abidal with the amrani/mariin phenotype.
-

- Afar woman with the amrani/mariin phenotype.
-

- Tuareg Berber woman with the amrani/mariin phenotype.
-

- Somali woman with the beidan/cad phenotype.
-

- Tuareg Berber woman with the beidan/cad phenotype.

Somali model Jawahir Ahmed (Darod clan), center, and North African contestants at a beauty pageant. Cushitic individuals with an cad phenotype have a similar complexion as North African individuals with a beidan phenotype, though they are often taller like the Tuareg Berbers.

Distribution of oblique eye folds and other forms of palpebral fissure (eye slits) among individuals in southern Somalia (Gallo (1979)).
Intermarrying with adjacent Arab or Persian groups is also clearly not responsible for these phenotypes. Gallo (1979) reports that most individuals in southern Somalia have either oblique eye folds (29.9%), slight Mongolian eye folds (19.9%), or external eye folds (19.9%), with the remainder possessing normal palpebral fissures/eye slits (36.6%). However, the southern Arabs generally do not have oblique eye folds nor are they particularly tall, and most are instead brachycephalic (broad-skulled). While Persians are frequently dolichocephalic (long-skulled; the cephalic/cranial index being a key difference between them and the southern Arabs), they too do not tend towards high stature or almond-shaped eyes (cf. Coon (1939)). (For more details, see Are you sure that the oblique eye folds, which are common among Afro-Asiatic speakers and Nubians in the Horn and North Africa, are not due to absorption of Khoisan peoples? What makes you ascribe this trait to Central Asian or East Asian admixture? and Is there any other evidence of an ancient Central Asian or East Asian-related presence in the Nile Valley?.)

Ethnic Somali women in northern Somalia. Almond-shaped eyes, which frequently characterize Berber individuals with the beidan phenotype, are also rather common among Cushitic individuals with the “pure” cad and mariin phenotypes. Ancient DNA analysis has confirmed that this similarity, like the resemblance between the mariin and amrani phenotypes, originates from the fact that the ancient Cushites were a population of North African ancestral stock.

Somali man with oblique eye folds. This form of palpebral fissure or eye slits likely evolved as an environmental adaptation among early Afro-Asiatic-speaking populations in the Sahara. Such eye folds are still rather frequent among Cushitic and especially Berber speakers; particularly individuals with the cad/beidan and mariin/amrani phenotypes.

Cephalic indices of populations in Northeast Africa, the Arabian Peninsula and Near East. The Cushitic and Ethiosemitic speakers and the North Arabs (Bedouins) are dolichocephalic (CI ≤74.9), whereas the South Arabs and Armenians are brachycephalic (CI ≥80).
Moreover, recent Arab and Persian admixture among Cushitic speakers in the Horn is primarily confined to the Benadiri Somalis (Reer Xamar or Xamar Cad-Cad), the latter of whom dwell in the coastal Benadir region in southern Somalia rather than in the northern parts of the country (Puntland and Somaliland territories) and other areas where cad individuals are concentrated. The early Benadiri intermarried with Arabian and Persian merchants and established the Somali-Arab-Persian ruling dynasties of the Mogadishu Sultanate. In this regard, the 1st century CE Periplus of the Erythraean Sea indicates that the ancient Azanians — the Cushitic ancestors of the Benadiri Somalis and the Bravanese, a subgroup of the Benadiri based in the port town of Brava — interbred with Arab traders on the coast of southern Somalia. This is why, to this day, especially tall Benadiri/Bravanese individuals can still be found (i.e., the great height was inherited from the Benadiri/Bravanese’s “giant” Cushitic Azanian progenitors, the latter of whom the Periplus describes as being “very great in stature”). Ibn Battuta also informs us that at the time of his visit in 1331, the Sultan of Mogadishu was one Abū Bakr ibn ‘Umar, a “Berber” originally from northern Somalia (Upshur et al. (2011); Freeman-Grenville (1963)). This first dynasty of the Mogadishu Sultanate, known as the Fakr ad-Din dynasty, was succeeded after 1500 by al-Muẓaffar dynasty (Freeman-Grenville (1963)). Correspondingly, Immel and Kleiber (2009) observed that most Benadiri individuals today carry the E1b1b and T paternal haplogroups like other ethnic Somalis (~52%; 35.48% E1b1b and 16.13% T), albeit with notable frequencies of the Arab-mediated J clade (~29%) and the Persian-affiliated R lineage (~6%). Genome analysis of modern Benadiris similarly indicates that these individuals mostly bear Cushitic-related ancestry (averaging ~50%), with significant western Asian admixture derived from peninsular Arabs (~25%) (see details on Who were the ancient Azanians?). Benadiri (as Arabized Cushites) are thus the Horn of Africa’s equivalent to North Africa’s Arabic-speaking populations (Arabized Berbers).

Admixture analysis of a Bajuni individual of Swahili type from Somalia. The person (labeled S25) was found to mainly harbor Bantu-related ancestry (light green component) with minor West Eurasian admixture, like other Swahili speakers from the Comoros and East Africa. However, unlike the Comorian Swahilis, the West Eurasian admixture borne by this Bajuni individual and the other East Africa Swahili samples is primarily associated with Cushitic-speaking populations of the Horn region (dark grey component) rather than Arabs (red component) or Iranians (light pink component) (Brucato et al. (2019)).
Benadiri/Bravanese are also ancestrally distinct from the Swahili. Although the Bravanese recently adopted the Swahili language during the Zanzibar Sultanate’s presence in Benadir, other Benadiri still speak their native Cushitic tongue (Somali). By contrast, the Swahili are people of Bantu origin and language, who have some Cushitic, Arab and Persian admixture; particularly among the Bajuni Swahili in southern Somalia. Brucato et al. (2018) observed that Swahili Bantu individuals of coastal East Africa (Mombasa, Lamu and Kilifi in Kenya) and Swahili Bantu individuals of the Comoros Islands predominantly bear Bantu-related autosomal DNA (~75%), with some minor Eurasian admixture derived from absorbed Cushitic peoples and Austronesian, Arab & Iranian peoples, respectively. Raaum et al. (2017) likewise report that the Swahili carry diverse Y-DNA haplogroups (including the Arabian-mediated J clade and the Cushitic-affiliated E1b1b and T lineages), though their most frequently occurring paternal haplogroup is the Bantu-associated E1b1a clade. The mtDNA of both the early Swahili (74%-89%) and modern Swahili (93%) almost entirely consists of African derivatives of the macrohaplogroup L, which are shared with neighboring Bantu and Nilotic populations (also see Msaidie et al. (2011)). Lastly, ancient DNA analysis of Swahili individuals at various Middle Ages coastal sites in Kenya and Tanzania indicates that they sustained considerably more gene flow from western Asia than their comparatively less admixed modern Swahili Bantu successors (50% Near Eastern-related admixture for the former vs. under 25% for the latter). The west Asian admixture that these medieval Swahili specimens carry is also primarily associated with Persians, whereas contemporary Benadiri/Bravanese Somalis to the north mostly derive their west Asian admixture from peninsular Arabs (cf. Brielle et al. (2022), Proctor et al. (2015), and Sloan (2010) for more ancient DNA of the early Swahili Bantus.)
-

- Sultan Bihi Mumin (Darod Somali, Cushite). Notice the great height, which is common among Cushitic individuals of similar cad physiognomy.
-

- Sef bin Ahmed, brother of the ex-Governor of Mogadishu (Benadiri Somali, Arabized Cushite).
-

- Arabic-speaking Libyan man of Cyrenaica (Arabized Berber).
-
- Bajuni man of Cushitic type (Katwa clan, assimilated Garre Somali origin).
-

- Bajuni man of Arab type (Nufayl clan, assimilated Omani origin).
-

- Bajuni man of Swahili type (Arab & Cushitic-admixed Bantu).
-

- Swahili man (slightly Arab & Cushitic-admixed Bantu).
-

- Swahili woman (slightly Arab & Cushitic-admixed Bantu).
Additional evidence that the mariin and cad phenotypes were not introduced by Arabs or Persians comes from historical texts. For example, in the 16th century Andalusian explorer Leo Africanus informs us that “the people of Adel are of the colour of an olive.” This is an allusion to the denizens of the Adal Sultanate, which he describes as “a very large kindome, and extendeth from the mouth of the Arabian gulfe to the cape of Guardafu.” The Portuguese missionary Francisco Alvares, a contemporary of Leo Africanus, confirms that indeed “the kingdom of Adel (as they say) is a big kingdom, and it extends over the Cape of Guardafuy, and in that part another rules subject to it.” Correspondingly, this realm is the exact general area in northern Somalia (Puntland and Somaliland), Djibouti and eastern Ethiopia where Cushitic individuals with the cad phenotype are still concentrated. According to the chronicler Shihāb al-Dīn’s coeval treatise Futuh Al-Habasa (“Conquest of Abyssinia”), the Adalites largely consisted of ethnic Somali clans, among whom were: the Gorgorāh Dir; Harti Darod (Warsangali, Majerteen, Dhulbahante), whom he calls the “people of Mait” after the northwestern port city of Mait; Marraiḥān Darod; Girri Ogaden Darod; Yabarre Ogaden Darod; Hawiya; and Habr Maqdi Isaaq.
(*N.B. Leo Africanus asserts that the Adalites of the Horn were of Arabian origin. In connection with this claim, modern genome analysis of ethnic Somali individuals born in the northeastern Puntland region of Somalia, who mostly belong to the Majerteen subclan of the Darod, has detected a predominant non-African ancestry. This Eurasian ancestry is comparable in frequency to that carried by North African individuals. Following Clark (1954), we suggest that this is because these northern Somali individuals are of relatively pure Cushitic stock rather than of recent peninsular Arab origin. For details, refer to Which Afro-Asiatic-speaking population of the Horn region has the most non-African ancestry?. Also see I have heard that the V12 and V32 subclades of the paternal haplogroup E1b1b, which are today common among Afro-Asiatic speakers in Northeast Africa, are old Arabian lineages. Is this true?.)
-

- Stone ruins belonging to the medieval Adal Sultanate in Zeila, northern Somalia. According to Leo Africanus, the polity was “a very large kindome, and extendeth from the mouth of the Arabian gulfe to the cape of Guardafu.” The explorer also indicates that “the people of Adel are of the colour of an olive.” This is an allusion to the olive-skinned cad phenotype that is found among the Cushitic inhabitants of the Horn of Africa.
-

- The earliest known map of the whole continent of Africa, published while the Adal Sultanate was still extant, identifies the kingdom as the “Regnum Seylam” after its political seat Zeila. As also confirmed by Leo Africanus and Francisco Alvares, the territory is shown to have occupied an area in the Horn region stretching from present-day Djibouti in the northwest to Cape Guardafui in the northeastern Puntland region of Somalia (Münster (1554)).
-

- Somali general Abdullahi Ahmed Irro (Darod clan), with an example of the cad phenotype among Cushitic individuals in Puntland, northeastern Somalia, the eastern limit of the former Adal Sultanate.
-

- Somali merchant Omar Musa (Isaaq clan), with an example of the cad phenotype among Cushitic individuals in Somaliland, northwestern Somalia, the central part of the former Adal Sultanate.
Oral testimonies transcribed by colonial officials in British East Africa during the early 20th century are in a similar vein. One such recorded eyewitness account from a Karimojong Nilote elder describes a cattle raid in the late 1800s that his tribe had made against one of the last remaining Cushitic groups in Kenya (which has since been assimilated into a hunter-gatherer community called the Oropom). He avers that this pastoralist population was “very light skinned and had pronounced slant eyes[…] The men wore their hair long in a pigtail, like Indian women” (cf. Wilson (1970)). Evidently, this is a description of the “white Galla,” whom J. C. Prichard and other early European adventurers reported having encountered in the area i.e., Cushitic individuals with the cad phenotype.
-
-

- A Somali woman with the cad phenotype. Cushitic individuals possessing this physiognomy are quite light-skinned (olive to light brown in tone) and often rather tall, usually with a slim build. They were known in the early colonial period as the “white Galla” due to their fair complexion.
-

- Somali entrepreneur Muna Magan (Isaaq clan), with a slightly swarthier example of the cad phenotype. We know from forensic analysis that the Caucasoid facial features, tall height, soft-textured hair, light complexion, slender build and almond-shaped eyes were all inherited from the early Cushites.
-

- Oromo women with another example of the cad phenotype. J. C. Prichard and other early European adventurers reported having encountered “white Galla” individuals near the Webi Shebelle river on the southern Ethiopia-Somalia border. Historiolinguistic analysis indicates that this area was occupied by various Cushitic groups, including Oromo speakers. Thus, the “white Galla” of yore were evidently Cushitic individuals with an cad physiognomy.
-

- An Afar woman showing another example of the cad (“olive-skinned”) phenotype, which is found among the Cushitic peoples. Genome analysis suggests that among the Afro-Asiatic speakers from the Horn region, alleles for light pigmentation were derived from two West Eurasian sources (European Steppe and Anatolian Neolithic components) and an East Eurasian source (ancient East Asian component). For details, see Genetic affinities of the Cushitic, Ethiosemitic and North Omotic-speaking populations of the Horn of Africa).
-

- A Hadiya Sidamo girl with an example of the cad phenotype among Cushitic children. As with all of the other people in this gallery, this little girl was born to two ethnically Cushitic parents. Her physiognomy is not due to recent intermixture with non-Cushitic peoples.
-

- A Somali teenage girl with an example of the cad phenotype among Cushitic adolescents.
-

- Somali model Jawahir Ahmed with the cad phenotype. Like Muna Magan, she is an ethnic Somali. Both of Jawahir’s parents belong to the Majerteen Darod clan, centered in the northeastern Puntland region of Somalia.
-
![Berber dancer and actress Sofia Boutella. Note the close likeness to Jawahir Ahmed. This calls to mind Coon (1939)'s assertion that "the Tuareg in their pure form belong to a specialized Mediterranean sub-type[...] They resemble the East African Hamites very closely, and especially the whiter element among the Somali."](https://landofpunt.files.wordpress.com/2015/11/berber-beidan.jpg?w=209&h=300)
- Berber dancer and actress Sofia Boutella. Note the close likeness to Jawahir Ahmed. This calls to mind Coon (1939)‘s assertion that “the Tuareg in their pure form belong to a specialized Mediterranean sub-type[…] They resemble the East African Hamites very closely, and especially the whiter element among the Somali.”
-

- A Wolayta woman with an example of the cad phenotype found among North Omotic speakers.
-

- An Oromo man with an example of the cad phenotype among Cushitic males.
-
-









-

- Inner coffin of the Kushite princess Amenirdis I, 25th Dynasty (PAHMA).
-

- Illustration of the Kushite princess Amenirdis I, depicted with an cad phenotype (Brunton (1929)).
-

- An Oromo woman with the cad phenotype. Based on her inner coffin, the Kushite princess Amenirdis I probably would have had a similar physiognomy.
-

- An ethnic Somali woman at the maximal end of the cad phenotype spectrum. Although it is doubtful that the Kushite princess Amenirdis I was this pale, ancient Egyptian artwork does suggest that there were many peoples with a very fair complexion who inhabited the Nile Valley.

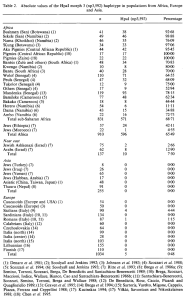




















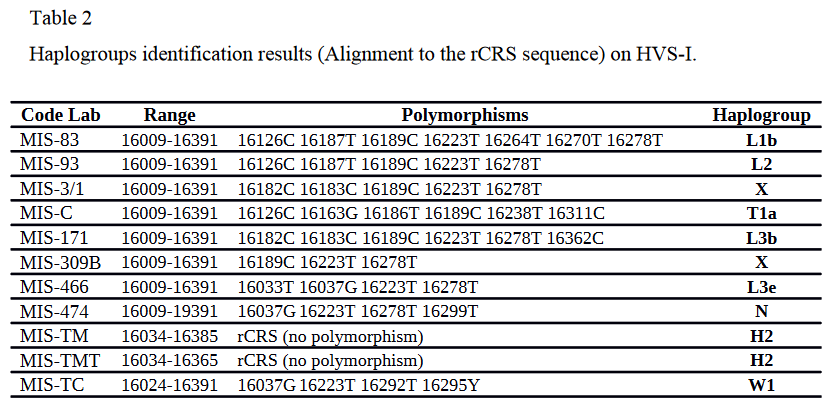






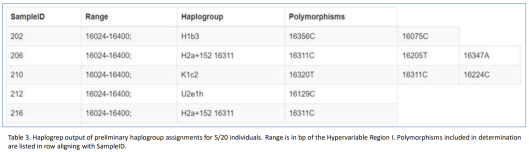
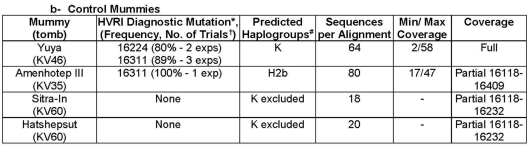

















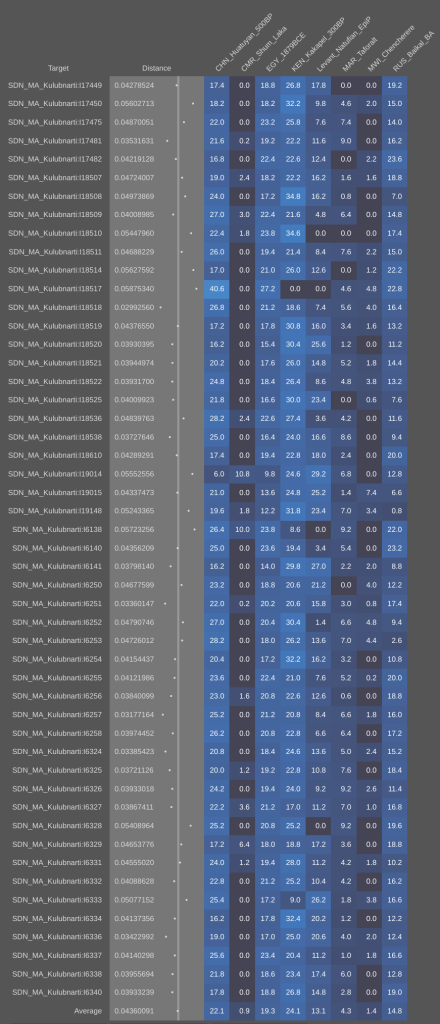
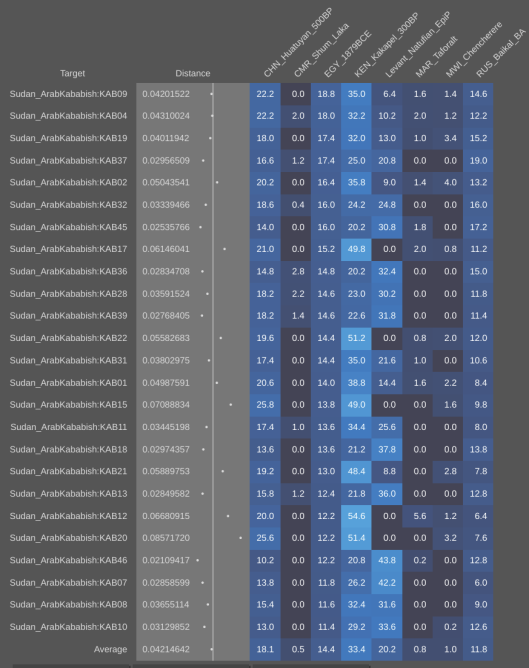




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}